1.a. AAbb × aaBb b.Aabb × aaBb c.AaBB × aabb
2.a. t1t1t2t2;b. T1t1T2t2;c.除了t1t1t2t2外的任一个基因型,即T1_ T2_,T1_ t2t2,t1t1T2_。
3.a. 1)GGWxwx 2)GGwxwx 3)ggWxwx 4)ggWxWx 5)GGWxWx 6)GgWxWx 7)Ggwxwx 8)GgWxwx
b.
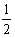 gWx;
gWx;
gwx
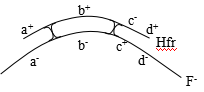
c.
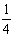 GWx;
GWx;
Gwx;
gWx;
gwx
4.a.从第一种杂交知紫花为显性,从第二种杂交知高茎为显性。
b.两种杂交产生的后代是:
PPtt × pptt → Pptt(紫花矮茎)
ppTT × pptt → ppTt(白花高茎)
这两种后代相互杂交,第三代是:
Pptt x ppTt t →
PpTt,
Pptt,
tpTt,
pptt。
所以第三代有比例相等的紫花高茎、紫花矮茎、白花高茎、白花矮茎四种类型。
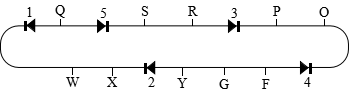
5.a. P ffcc × FFCC
↓
F1 FfCc
b.54季节性开花、皱叶,18连续开花、皱叶,18季节性开花、非皱叶,6连续开花、非皱叶。
c. 24
d. 12
e. 6
6.因为 AABB × aabb
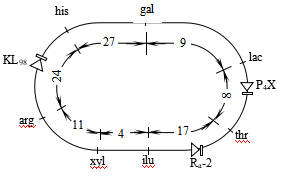
↓
AaBb
所以该黑狗的基因型是AaBb。这样,它与同基因型狗交配,子代的情形是:
AaBb × AaBb
↓
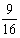 AABB
AABB
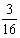 AAbb
AAbb
aaBB
 aabb
aabb
黑色 红色 赤褐色 柠檬色
7.两对具有互补的非等位基因控制花色遗传。这两对基因突变并自交纯合,形成CCpp或者ccPP的白色个体基因型。ccPP或CCpp × CCPP →紫花。ccPP x CCpp →紫花
8.a.设卷翅基因为L,正常翅基因为l;短刚毛基因为B,长刚毛基因为b。根据题意分析卷翅基因L和短刚毛基因B都是显性基因。
b.亲本的基因型为♀Llbb;♂llBb.
c.卷翅长刚毛(Llbb),卷翅短刚毛(LlBb),正常翅长刚毛(llbb),正常翅短刚毛(llBb)。
9.
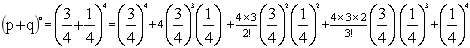
具有四个显性性状的个体为
 ,具有三个显性性状一个隐性性状的个体为
,具有三个显性性状一个隐性性状的个体为
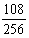 ,具有两个显性性状两个隐性性状的个体为
,具有两个显性性状两个隐性性状的个体为
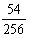 ,具有一个显性性状三个隐性性状的个体为
,具有一个显性性状三个隐性性状的个体为
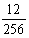 ,具有四个隐性性状的频率为
,具有四个隐性性状的频率为
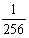 。
。
10.a. 3:1; b. 3:1; c. 1:2:1; d. 2:1:1; e. 1:1:1:1;
一、判断对错
1.√
2. √
3. √
二、选择正确答案
1.①b ②a ③c ④c ⑤d
2. c
三、填空
1.多因一效
2. 8;27;8
3.同一位点上的等位基因的相互作用
4.互补;积加;重叠;显性上位;隐性上位;抑制作用
5.1/64
6.(3:1)n;2n;(1:1)n;2n
7.基因内;基因间
8.分离;自由组合
四、名词解释
基因互作:不同对基因间的相互作用
互补作用:两种以上基因同时存在时,互相补充而表现另一种性状,这种基因间的相互作用称为互补作用。
互补基因:协同作用下产生某一性状的若干非等位基因,这些基因共同存在时才出现某一性状。
积加作用:两种以上显性基因同时存在时产生一种性状,单独存在时产生能分别表现相似的性状。
重叠作用:不同对基因互作时,对表现型产生想他的影响。
重叠基因:在重叠作用中表现相同作用的基因。
上位性、下位性:两对以上独立遗传基因共同对艺性状发生作用,而且其中一对基因对其他基因的表现有掩盖作用,这种情形称为上位性,后者被前者所掩盖,称为下位性。
显性上位作用:在两对以上互作的基因中,其中一对显性基因对其他对基因其上位作用。
隐性上位作用:在两对以上互作的基因中,其中一对隐性基因对其他对基因其上位作用。
抑制作用:在两对以上独立遗传的基因中,其中一对显性基因本身并不控制性状表现,但对其它对基因的表现有抑制效应。
基因内互作:同一位点上的等位基因的相互作用,表现为显性或不完全显性和隐性。
基因间互作:不同位点上的非等位基因间的相互作用。
多因一效:许多基因影响同一性状的表现。
一因多效:一个基因影响许多性状发育的现象。
返祖现象:隔若干代以后,出现于祖先相似性状的遗传现象。
第一章
连锁遗传规律
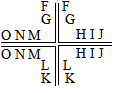
一、回答问题
1.475淡红眼,其中♀+car//+car,+car//Y♂各为1/2;475棒眼其中♀B+r//+car,B+//Y♂各为1/2;25正常眼,其中♀++//+car,++//Y♂各为1/2;25棒眼淡红眼,其中♀Bcar//+car,Bcar//Y♂各为1/2。
2.假设a和b间还有一基因o,则顺序为a-o-b,那么:
交换值(a-o)=重组体率(a-o)+双交换值……①
交换值(o-b)=重组体率(o-b)+双交换值……②
①+②得,交换值(a-b)=重组体率(a-b)+2×双交换值
20% =18%+2×双交换值
双交换值=(20%-18%)/2
=1%
3.TtHh×TrHh→T_H_,T _hh,ttH_,tthh,数目最少的子代的基因型应为重组体tthh。
所以,交换值=2×
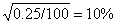 ,两基因之间的距离为10个遗传单位。
,两基因之间的距离为10个遗传单位。
4.AaEeFf可形成8种配子,A为独立,ef连锁,故Aef配子比例为
 。
。
5.实际双交换值为:0.19×0.2×0.6=0.02=2%
a-b间重组值=19%-2%=17%
b-d间重组值=20%-2%=18%
子代果蝇基因型及其数目:
双交换型:ABD/abd和abd/abd各为:

a-b重组型:ABd/abd和abD/abd各为:
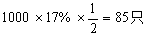
b-d重组型:Abd/abd和aBD/abd各为:

亲型:AbD/abd和aBd/abd各为:
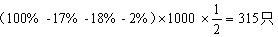
6.双交换值=0.2×0.3=0.06
各类型配子的比例为:
AbD和aBd各为0.06/2=0.03
Abd和aBD各为(0.3-0.06)/2=0.12
ABd和abD各为(0.2-0.06)/2=0.07
ABD和abd各为[1-2(0.03+0.12+0.07)]/2=0.28
7.连锁,因为与9:3:3:1相比,亲型多于理论值,重组型少于理论值。A与B间的距离为40%。
8.从理论上可以推知杂种F1形成4种配子,即Ab、AB、aB、ab。其中AB和ab为重组型。从实验结果可知,AB型配子所占比例为[68/(204+68) ×100%≈25%。那么ab型配子也应该为25%。4中配子比例相等,因此可断定ab间无连锁。
9.两基因连锁;有交换发生;交换值为16%;发生交换的性母细胞数位32%。
10.+bc和a++各为2.5%,ab+和++c各为5%,+++和abc各为42.5%。
11.a.三基因连锁;b. A与B相斥,A与C相引;c. b-a-c;d. b-a,30%;a-c,10%;e.无干扰。
12.a.方式交换的配子是AB和ab,未发生交换的配子为Ab和aB;
b.两基因间的遗传距离为16个图距单位;
c.减数分裂前期I非姊妹染色单体间发生交换的百分率为32%。
13.AB:Ab:aB:ab=(47+201):(43+207):(43+207):(47+205)≈1:1:1:1
AD:Ad:aD:ad=(47+207):(43+201):(43+205):(47+207)≈1:1:1:1
BD:Bb:bD:bd=(47+43):(201+207:(207+205):(43+47)≠1:1:1:1
所以只有BD两基因连锁,基因型为A/a Bd/bD。交换值(b-d)=[(90+90)/1000] ×100%=18%
14.有160个孢母细胞发生了单交换。
15.在杂种亲本产生的所有配子中,只有ABd类型配子测交后才能出现A_B_dd个体。其他类型的配子均不能产生次基因型的后代。如果三基因为独立遗传,杂种亲本必将产生8种比例相同的配子,ABd型配子的比例应为
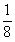 ,测交后的的A_B_bb的数字为
,测交后的的A_B_bb的数字为
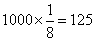 ,与结果不符。如果三基因中两基因连锁,另一对基因属独立遗传,杂种亲本产生的8类配子中将有两类比例值,没四种比例相同,而(330/1000)×4=1.32﹥1,与理论不符。所以唯一的可能是三基因连锁。
,与结果不符。如果三基因中两基因连锁,另一对基因属独立遗传,杂种亲本产生的8类配子中将有两类比例值,没四种比例相同,而(330/1000)×4=1.32﹥1,与理论不符。所以唯一的可能是三基因连锁。
16.如果两基因为独立遗传,那么双隐性个体应为
 ,而实际为16%。所以两对基因连锁。双隐性配子的比例为0.4,交换值=1-(0.4×2)=0.2=20%
,而实际为16%。所以两对基因连锁。双隐性配子的比例为0.4,交换值=1-(0.4×2)=0.2=20%
17.

 从测交子代的8中表现型的个体数看,比例为四多四少,每四种表现型比例一样,可知三对基因中有两对基因连锁,一对基因独立,从杂合体所成的亲型配子(ABC、ABc,abC、abc)看,A和B,a和b总是连在一起,与C(c)重组,所以可知AB连锁,C独立。AB间交换值为(20×4)/(20×4+200×4)=0.091。所以三基因在染色体上的位置是: A 9.1 B C
从测交子代的8中表现型的个体数看,比例为四多四少,每四种表现型比例一样,可知三对基因中有两对基因连锁,一对基因独立,从杂合体所成的亲型配子(ABC、ABc,abC、abc)看,A和B,a和b总是连在一起,与C(c)重组,所以可知AB连锁,C独立。AB间交换值为(20×4)/(20×4+200×4)=0.091。所以三基因在染色体上的位置是: A 9.1 B C
18.
 a.三基因的顺序为t-s-g; b. t 17.4 s 26.7 g c.符合系数=0.04/(0.174×0.267)=0.86
a.三基因的顺序为t-s-g; b. t 17.4 s 26.7 g c.符合系数=0.04/(0.174×0.267)=0.86
19.如果两基因为独立遗传或不完全连锁,自交F2应有重组类型出现。而实际上F2代中只有两种亲本类型,而且比例为3:1,由此可知两对基因连锁,而且为完全连锁。
20.从题中得知,aBc和AbC为双交换型配子,双交换率为0.05×0.1=0.005=5%,因此两种配子的频率各为0.25%。
21.实际双交换值=0.7×0.2×0.11=0.0154
a-b间重组率=0.2-0.0154=0.1846
b-d间重组率=0.11-0.0154=0.0946
ABD/abd和abd/abd个体的比例各为0.1846/2=9.23%
AbD/abd和aBd/abd个体的比例各为0.0946/2=4.73%
ABd/abd和abD/abd个体的比例各为0.0154/2=0.77%
Abd/abd和aBD/abd个体的比例各为(1-0.0154-0.1846-0.0946)/2=35.27%
22.当符合系数为1时:
双交换值=0.1×0.14×1=0.014
t-h间重组率=0.1-0.014=0.086
h-f间重组率=0.14-0.014=0.126
亲型比例=1-0.014-0.086-0.126=0.774
thf/thf的频率为0.774/2=0.387
当符合系数为0时,说明无交换发生,所以亲型的比率=1-0.1-0.14=0.76,thf/thf的频率为0.76/2=0.38
23.a.雌雄性别性状表现不同,断定属性染色体上基因遗传。
b.亲本的基因型及其遗传过程如下:
P: aBD//aBd×Abd//Y
↓
F1 ♂ aBD//Y×aBD//Abd♀
↓
F2 雄性 雌性
aBD//Y aBD//aBD
Abd//Y Abd //aBD
abd//Y abd //aBD
ABD//Y ABD //aBD
aBd//Y aBd //aBD
AbD//Y AbD //aBD
abD//Y abD //aBD
ABd//Y ABd //aBD
雄性个体中表现为8种表现型;雌性个体中仅有A_B_D和aaB_D_两种类型,而且比例相同。
c.只用雄性个体就可以估算出基因间距离:
a-b间距离:[(27+26)/300]+[(4+3)]=20%,为20个遗传单位。
b-d间距离:[(13+10)/300]+[(4+3)]=10%,为10个遗传单位。
24.在无干扰情况下,意味着两个单交换独立发生,则有:
∵双交换值=AB间交换值×BC间交换值
∴BC间交换值=双交换值/AB间交换值=(2×0.0025)/0.05=0.1
25.a. AaBbb产生4种配子,两多两少,所以可知AB连锁,aB和Ab为亲型配子。
b. AB间交换值为(18+17)/(18+17+34+31)×100%=35%,图距为35个遗传单位。
c.因为aB和Ab为亲型配子,所以AaBbb的两个亲本基因型为aB//Ab和Ab//Ab。
26.第一个杂交组合中高圆的基因型为TP//tp,第二个杂交组合中高圆的基因型为Tp//tP。后代个表现的比例为:54高茎圆果:21高茎梨形果:21矮茎圆果:4矮茎梨形果。
27.因为dsmp/dsmp占后代群体的4/100,所以子代群体至少应种x=(100×20)/4=500株。
28.依题意知两基因间交换值为8.6%。F2出现裸粒散穗数目标性状个体比例为4.1%。非目标性状个体所占比例接近96%。
29.a. 8种类型配子的比率为EDR20.5%,EDr20.5%,edR20.5%,edr20.5%,EdR4.5%,Edr4.5%,eDR4.5%,eDr4.5%。
b.稳定的矮秆、抗锈、正常叶植株即基因型为ddRREE的植株,其出现期望百分率为(dRE)2=(4.5%)2≈0.2%
30.a.母本植株籽粒果皮无色,胚乳饱满,非糯。
b.F1植株表现抗大斑病。
c. F1果穗籽粒全为巧克力色。胚乳分离为饱满非糯质15%,饱满糯质35%;凹陷非糯质35%;凹陷糯质15%。抗病与不抗病比例为1:1.
31.R1R2R320% R1R2r32 0% r1r2R320% r1r2r320% R1r2R35% R1r2r35% r1R2R35% r1R2r35%
32.亲本基因型:♀Aa×♂aa;有角A为显性,但只在♂中表现,在♀不表现,属从性遗传。
33.a.14.5%;b.25%;c.4%
34.交换值=(交换子囊数/子囊总数)×100%×1/2=[(2+3+2+1)/(48+44+2+3+2+1)] ×100%×1/2=4%,基因a与着丝点间的距离为4个遗传单位。
35.雄性亲本基因型X+YJj,雌性亲本基因型X+XpJj。
36.♀:XaXb花色 ♂:XbY橙黄色
37.基因a与着丝点的距离为
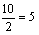 ;基因b与着丝点的距离为
;基因b与着丝点的距离为
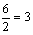 。如果两基因为着丝点的同侧,那么它们之间的距离为5-3=2。.重组型孢子占2%,AB型孢子为1%。如果两基因位于着丝点两边,那么它们之间的距离为5+3=8,重组型孢子占8%,AB型孢子为4%。
。如果两基因为着丝点的同侧,那么它们之间的距离为5-3=2。.重组型孢子占2%,AB型孢子为1%。如果两基因位于着丝点两边,那么它们之间的距离为5+3=8,重组型孢子占8%,AB型孢子为4%。
38.遗传图上的数字是累加的,而且仅列出了某些基因。
39.母亲XCXc;父亲XCY;孩子XcO。
40.人类男性性染色体为XY,女性性染色体为XX。男性可产生含X和Y染色体两类数目相等的配子,而女性只产生一种含X染色体的配子。精卵配子结合后产生含XY和XX两类比例相同的合子,分别发育成男性和女性。因此,男女性比接近1:1。
41.在雄性群体中两者以相同的频率出现,在雌性群体中显性伴性基因表现出来的频率高。
42.尽管这只母鸡发生了性反转,但它的性染色体并未发生改变,仍为ZW型,因此将有下列遗传过程:
性反转的公鸡ZW×ZW正常母鸡
↓
ZZ ZW ZW WW(死亡)
后代性别比例为2雌:1雄。
43.如果母亲的两条性染色体上都有携带色盲基因,那么:XcXc×XCY→XCXc,XcY,他们的女儿全部正常,儿子全部色盲。
如果母亲只有一条染色体上基因正常,那么:XCXc×XCY→XCXC,XCY,XCXc,XcY,他们的女儿全部正常,儿子中将有一半可能为色盲。
44.该基因位于Y染色体上。
45.雌果蝇的一条性染色体含有隐性致死基因。
46.亲代基因型 XCXc × XCY
↓
子代基因型 XcY XCXC XCXc
↓ ↓ ↓
孙代基因型 XCY XCY XCY,XcY
47.男人24;女人23;雌蝗虫中12。
48.a.女儿全部正常,儿子有一半可能为色盲; b.无论女儿和儿子各有一半可能为色盲。
49.限性遗传。
50.蜜蜂的正常受精卵(2n)发育成雌性,孤雌生殖发育的单倍体(n)为雄性,如果用a表示黄绿色的隐性基因,用A表示正常的褐眼基因;
♀aa×♂A
↓
孤雌生殖a、正常受精Aa
后代中雌蜂全部为正常褐眼,雄峰为黄绿色眼。
51.伴性遗传。隐性色盲基因在X染色体上,Y不带基因,XY为男性性染色体组成。
52.同源染色体在减数分裂前期I非姊妹染色单体的交换。相邻两次交换相互影响和抑制的现象交染色体干涉。实际双交换值与理论双交换值之比称符合系数。符合系数为1时,表明两次单交换独立发生;为0时,无双交换发生。符合系数范围为0-1,越大说明双交换发生的可能性大,反之亦然。
53.儿子XXY和XY各占一半;女儿XXX和XX各占一半。
54.ZbW;ZBZB或ZBZb;ZBW;ZbZb。
55.分离规律是遗传的最基本规律,是研究任何一对等位基因所控制的性状的传递规律。
独立分配规律和连锁遗传规律都是研究两对或两对以上等位基因锁控制性状的传递规律。区别在于独立分配规律是研究位于非同源染色体上的基因所控制的性状共同遗传时的规律,而连锁遗传则是研究位于同一染色体上不同基因所控制性状共同遗传时的规律。但是,独立或连锁中的任何一对基因所控制的性状仍受分离规律的支配。
56.a. 5%; b. 9.8%
57.玉米、蜜蜂为同配性别;人、鸡为异配性别。
58.a.两亲本基因型为♀XNXn,♂XNY; b.细叶后代所占比例为1/4; c.细叶植株为雄性。
59.

 A 7.4 B 2.9 C ; A 4.5 C 2.9 B
A 7.4 B 2.9 C ; A 4.5 C 2.9 B
60.发生交换的孢母细胞所形成的配子中,只有半数属交换型配子。
61.XcXc。
62.很近。
63.20%。
64.a.连锁遗传,交换值为16%,重组型配子为ab和AB; b. AAbb和aaBB
65.
 着丝点 ← 25 → P
着丝点 ← 25 → P
66.白眼基因和桃红眼基因互为等位基因;白眼基因与朱红眼基因互为非等位基因。
67.a. ++//sp; b. ++ 48%;sp 48%;+p 2%;s+ 2%。
68.雌鼠的一条X染色体上含有隐性致死基因。
69.杂合体为雄亲,纯合体为雌亲。因为雄果蝇不发生交换,所以产生的朱红色和残翅之比为1:1。
70.22%。
71.a. Rt和rT; b.RT和rt。
72.
 基因顺序为:a d b
基因顺序为:a d b
73.两基因为完全连锁,绿苗亲本的基因型为Ab/aB。
74.女儿正常,儿子中有一半的可能为色盲。
75.雌性:雄性=2:1。
76.将基因型BabaTsts植株自交:
BabaTsts
↓

9Ba_Ts_ : 3Ba_tsts : 3babaTs_ : 1babatsts
正常雌性同株 雌株 雄株 仅顶端有雌花
自交后代中选择基因型为babaTs__和babatsts的植株连续杂交几代后,就可将玉米变成雌雄异株植物。
77.交叉现象的发生表明非姊妹染色单体间已发生了交换,而发生交换的孢母细胞所形成的四个配子中,只有两个是重组型的,另两个仍是亲型配子。就是说发生交换的孢母细胞所形成的配子中只有一半属交换型配子。因此重组率为交叉频率的1/2.
78.如果雌性果蝇的X染色体上有隐性(+→﹣)突变发生,可利用杂交后代雄性个体中隐性表型的比例来测定。如果雄性果蝇的X染色体上有隐性突变发生,可利用CIB测定法进行检测。
79.a. EeGgHh;b. EGH和egh;c. EGH : EGh : egH : egh = 1:1:1:1
80.遗传基因的区别:伴性遗传是指X染色体上或Z染色体上基因所控制的性状的遗传;限性遗传是Y染色体上或W染色体上的基因所控制的性状的遗传:从性遗传是指染色体基因所控制的性状在表现上受个体性别影响的现象。
性状表现的区别:伴性遗传的性状既可在雄性个体中表现,也可在雌性个体中表现;限性遗传的性状只限于在一种性别上表现;从性遗传的性状只出现于雄性或雌性一方,或一方为显性,另一方为隐性。
81.交换值即为重组型配子占总配子数的百分比。可通过测交法或自交发求之。
测交法:交换值=(重组型个体/个体总数)×100%
自交法:如果杂交亲本为相引相‘
交换值=

如果杂交亲本为相斥相
交换值=
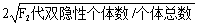
交换值、连锁强度和基因间距离的关系为:交换值越大,基因间的连锁强度越弱,它们之间的距离越远。反之,交换值越小,基因间的连锁强度越强,它们之间的距离越近。
82.设人类白化病可由不同的隐性基因决定。这对白化病夫妇中的一个是由纯合隐性基因型aa决定,另一个是由纯合隐性基因型bb决定的。它们的基因型分别为aaBB和AAbb。它们的子代基因型AaBb。由于基因A和基因B有互补作用,所以表现出正常。
83.色盲是由于染色体X上隐性基因控制的。男性性染色体组成为XY型,只需X染色体上携带色盲基因,就会表现出色盲性状。而女性X有两条染色体。如果一条性染色体携带色盲基因,并不表现色盲症状,只有两条X染色体同时携带色盲基因,才会表现出色盲症状。所以男性比女性更容易患色盲病。
84.a.同9:3:3:1的独立遗传比例相比较,F2四种表现型中亲本组合性状的实际数多于理论数,重组性状的实际数少于理论数。
b.同一染色体上的两个基因在某些性母细胞中可能发生非姊妹染色单体交换,有的则未发生交换。发生交换的可形成半数重组配子,半数亲型配子。而未发生交换的,只能形成亲型配子,所以重组率总是低于50%。
85.同源染色体的的异质等位基因有可能通过非姊妹染色单体的交换而互换位置,使姊妹染色单体异质化。只有到细胞第二次分裂,姊妹染色单体分离时,这对异质基因才能分离。
86.根据题意,只有aabbcc不表现表现,所以:
a. F1(AaBbCc)形成8种配子,abc配子占20%,自交后代中aabbcc的比例为(20%)2=4%,96%为Z表现型。
b.测交后代中aabbcc的比例为20%,Z表现性型为80%。
87.1)根据测交结果找出亲型和双亲交换型。
2)按双交换原理(亲型和双亲交换型基因型就是位于中间基因的对调)比较亲型和双亲交换型,找出双交换配子中哪个脱离了亲型配子的连锁群,则哪个基因就在中间。由此可以决定基因顺序。
3)计算两个交换值(必须加上双交换值)。
4)根据所得基因顺序和交换值绘制连锁图。
88.a.性染色体决定性别:其中有雄杂合型,即雄性的染色体为XY型,雌性的为XX型,如牛、马、羊等。还有雌性杂合型,即雌性性染色体为ZW型,雄性为ZZ型,如鸟类、蛾等。在雄杂合型中还有一XO种型,雌性个体有两条X染色体,雄性只有一条X染色体,故称XO型,如蝗虫、蟋蟀等。
b.染色体的倍数性决定性别:如蜜蜂、蚂蚁。正常的二倍体(2n)为雌性,孤雌生殖而来的单倍体(n)为雄性。
c.性染色体(X)和常染色体组(A)比例决定性别:如果蝇,当性染色体与常染色体比例(X/A)为1.0时,发育成雌性;0.5时为雄性;1.5时为超雌性;0.3时为超雄性;0.67时为中间性。
d.基因决定性别:如玉米的雌花序和雄花序分别受两对基因BaBa,TsTs所控制。
e.环境条件的影响:如黄瓜,苗期多施氮肥,适当缩短光照可提高雌花的数量。
89.益处:可使同源染色体的非姊妹染色单体间的基因重组,结果产生多种多样的配子,为变异提供了遗传的物质基础。
害处:打破了有益基因间的连锁。
90.雄果蝇的X染色体上载有控制小眼性状的一个显性基因B。其遗传过程如下:
♂ XBY× ♀XbXb
↓
♀ 65小眼XBXb ♂67正常眼XbY
91.基因型AaBb产生的配子种类和比例为:1 AB:1Ab:1aB:1ab; 基因型BbCc产生的配子种类和比例为:
0.45BC:0.05Bc:0.05bC:0.45bc;
基因型AaBbCc产生的配子种类和比例为:
0.225ABC:0.025ABc:0.025AbC:0.225Abc
0.225aBC:0.025aBc:0.025abC:0.225abc
92. 父亲AaBb×母亲Aabb
↓
1AABb:2AaBb:1AAbb:2Aabb:1aaBb:aabb
儿子中,3秃头褐眼:3秃头蓝眼:1非秃头褐眼:1非秃头蓝眼
女儿中,1秃头褐眼:1秃头蓝眼:3非秃头褐眼:3非秃头蓝眼
93.高株,红花
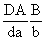 ×
×
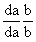 矮株,白花
矮株,白花
↓
高株、红花22.5% 高株、白花27.5%
矮株、红花2.5% 矮株、白花47.5%
94.a.朱红眼基因(v)位于X染色体上;
b.正交: ♂ XvY × ♀ X+X+
↓
♂X+Y暗红眼♀X+Xv暗红眼
反交: ♂ X+Y ×♀ XvXv
↓
♂XvY朱红眼♀X+Xv暗红眼
c. ♀X+Xv×♂XvY
↓
X+Xv暗红眼X+Y暗红眼XvXv朱红眼XvY朱红眼
95. 第一代♀:♂=1:1,第二代♀:♂=2:1
96.
 a. a + + b + + a b
a. a + + b + + a b
 b. a + + b
b. a + + b
c. a + + b + + a b
97.设色盲基因c,蓝眼基因b,母亲的基因为BbX+Xc,父亲的基因型为BbX+Y。
98.

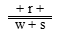
 在 与 的测交后代中,只有三基因w r s排列,后代中双交换型才为 和
在 与 的测交后代中,只有三基因w r s排列,后代中双交换型才为 和
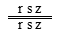
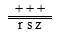 。在 与 的测交后代中,只有三基因r s z排列,后代中双交换型才为
。在 与 的测交后代中,只有三基因r s z排列,后代中双交换型才为

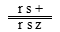 和 。所以基因在染色体上的位置为w r z s。
和 。所以基因在染色体上的位置为w r z s。
99.a. 父亲(aa或Aa)×母亲秃顶(AA)
↓
非秃顶男人(aa)×非秃顶女人(Aa)
↓
Aa aa
女儿全为正常,儿子中一半为秃顶
b.非秃顶男人(aa) ×秃顶女人(AA)
↓
Aa
女儿为正常,儿子全为秃顶
c.非秃顶父亲(aa)×母亲(AA或Aa)
↓
秃顶男人(Aa)× 非秃顶女人(aa)
↓
Aa aa
女儿全为正常,儿子中一半为秃顶
100.对同载在一条染色体上的基因群中的基因间关系和传递规律进行研究,对植物育种有一定指导意义。基因间的连锁关系研究得越清楚,越能提高育种的预见性。
连锁基因通过交换,使基因重新组合。根据交换值得大小,预计重组目标个体出现的频率,从而可适当安排杂交后代群体大小。
同载于一条染色体上的基因所控制的性状可表现一定的相关性。可根据这种相关性,选择某一个性状以达到选择另一个性状的目的。
二、判断对错
1.×;某些内外因素,如性别、年龄、温度等,可以影响交换值的变化。
2.×; 连锁群与染色体对数一致。
3.×; 也可能是独立遗传。
4.4. ×;Y染色体带有一些基因,两者可以配对。
5.×;两对连锁,另一对为独立遗传。
6.√
7.√
8.√
9.×;由异染色质和常染色质组成。
10.×;为相斥组。
11.×;这是从性遗传。
12.√
13.√
14.×;Y染色体上携带表现限性遗传的基因。
15.×;发生交换的性母细胞为20%。
16.×;在独立遗传时,双隐性个体所占频率也应为6.25%。
17.√
三、选择正确答案
1.b
2.b
3.c
4.1)c;2)c;3)b;4)d;5)a
四、填空
1.基因;性染色体;染色体的倍数性
2.40%;10%;20%
3.摩尔根
4.越弱;完全
5.XY;XO;ZZ;单倍体(n)
6.1
7.8;6
8.XXY;XO
9.少
10.弱;大
11.超雄
12.贝特生;摩尔根
13.性别;年龄;温度
14.加上双交换值
15.XX+AA
16.0~1;独立
17.A;B
18.12
19.6;2;4
20.基因定位;杂交;测交;三对
21.同一;1;第一次分裂的前;非姊妹染色单体片断的交换
五、名词解释
连锁遗传:位于同一染色体上的基因,作为一个行动单位而共同传递到一个配子中的现象。
相引相:甲乙两个显性性状联系在一起遗传、甲乙两个隐性性状联系在一起遗传的杂交组合。
相斥相:甲显性性状和乙隐性性状联系在一起遗传、乙显性性状和甲隐性性状联系在一起遗传的杂交组合。
连锁群:位于同一染色体上一组连锁在一起的许许多多基因。
完全连锁:一对染色体上的基因紧密连锁,完全不发生交换,测交后代中只出现亲本类型。
不完全连锁:一对染色体上的基因间或多或少地发生交换,测交后代中大部分为亲本类型,少部分为重组类型。
交换值:即重组率,指重组型配子数占总配子数的百分率。
遗传距离:以交换值表示两个基因在同一染色体上的相对距离。
基因定位:确定基因在染色体上的排列顺序和彼此间的距离。
连锁遗传图:即遗传学图。把一个连锁群的各个基因的排列顺序和相对距离标志在染色体上,绘制成图。
两点测验:以两对基因为基本单位计算重组率,从而确定基因在染色体上位置的遗传学方法。
三点测验:以三对基因为基本单位计算重组率,从而确定基因在染色体上位置的遗传学方法。
单交换:基因间分别发生的单个交换。
双交换:在一个四联体中发生的两个交换。
干扰:一个单交换发生后,影响其邻近位点再发生交换的现象。
符合系数:表示干扰程度的数学度量值。即

伴性遗传(性连锁):指性染色体上基因所控制的某些性状总是伴随性别而遗传的现象。
性染色体:与决定生物性别有关的染色体,携带专一基因,能部分决定个体性别的染色体。
常染色体:在生物许多成对的染色体中,除性染色体以外的其余各对染色体。
交叉遗传:子代与其亲代在性别和性状上出现相反表现的现象。
限性遗传:指某些性状只局限于雄性或雌性上表现的现象。
从性遗传:指位于常染色体的基因所控制的性状,由于内分泌及其他关系,使某些性状只出现于雌雄一方,或在一方为显性,另一方为隐性的现象。
X染色体:雄性是异配性别的生物的性染色体之一,在同配性别中有两个,异配性别中有一个。
Y染色体:雄性是异配性别的生物中,雄性个体细胞内与X染色体相配对的那条性染色体。
Z染色体:雌性是异配性别的生物的性染色体之一,在异配性别中有一个Z染色体,在同配性别中有两个。
四分子分析:对减数分裂所产生的四个单倍体细胞(四分子)所作的遗传分析,这种分析只能在各个四分子不相混杂的生物中才能进行,如担子菌、子囊菌等。
第二章
数量性状的遗传
一、回答问题
1.如果两杂交亲本的遗传组成不是极端类型,杂交F2代就有可能出现超亲现象,反之则无。例如:
A1A1A2A2a3a3×a1a1a2a2A3A3
↓
A1a1A2a2A3a3
↓
A1A1A2A2A3A3,……,a1a1a2a2a3a3
A1A1A2A2A3A3或a1a1a2a2a3a3对性状的作用就可能超过亲本。
2.两亲本应具有互补的高度基因。如:A1A1a2a2×a1a1A2A2
↓
A1a1A2a2
↓
A1A1A2A2。。。。。。。。。,a1a1a2a2
A1A1A2A2或a1a1a2a2的高度可能超过亲本,且稳定遗传。
3.a.利用基因型纯和的亲本估算:因为作亲本的都是纯种,遗传组成一致,所以基因型方差为零。就是说两亲本各自的表现型的差异完全是由环境引起的,所以亲本的表型方差就是环境方差,即

b.利用F1代群体估算:因为杂交亲本是纯合的,杂交F1代群体基因是一致的。因此可以认为F1代的表现型的差异也是来自环境的差异,及环境方差就等于F1的表现型方差
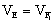 。
。
c.利用两亲本表型方差和F1的表型方差合计来估算:
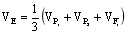
4.这种籽粒见到阳光才变红,说明它是由遗传和环境因素共同决定的。
5.数量性状呈连续性变异,而质量性状呈不连续变异;数量性状容易受环境条件的影响产生不遗传的变异,而质量性状不易受外界条件的影响;数量性状是受微效多基因控制的,而质量性状是受主基因控制。
由于数量性状表现为连续性的变异,因此研究其遗传动态只能采用统计学方法。而质量性状,由于呈不连续变异,可在杂种后代分离群体中按性状之间的差异将其归类分组,求出不同类型的比例,进而研究其遗传规律。
6.第一,数量性状的变异表现为连续的,杂种后代的分离世代不能明确分组。第二,数量性状一般容易受环境条件的影响而发生变异,这种变异一般是不遗传的。
7.遗传方差占总方差的比值为广义遗传力,而狭义遗传力是遗传方差中的加性方差占总方差的比值。广义遗传力的估算方法简便,但比较粗放。狭义遗传力剔除了广义遗传力中那些次要的、不稳定的遗传成分,因此数值要比广义遗传力小,但结果更加可靠。
8.花斑性状受主效基因和修饰基因共同控制,修饰基因数量的不同,造成个体间的这种差异。
9.所选的优良个体的优良性状是由于环境条件的差异引起的,属不遗传的优良性状。
10.a.父亲AABBCCDD,母亲aabbccdd;b.仅表现绿色眼睛。
11.亲本的基因型可能有多种形式,但两者必须呈互补状态,如A1A1A2A2a3a3a4a4×a1a1a2a2A3A3A4A4,每个A基因对表型的效应相同,每个a基因对表型的效应也相同。
12.甲AAbbcc,乙aaBbCc。
13.a. 155厘米的有Aabbcc,aaBbcc,aabbCc。
b.175厘米的有AaBBCC,AABbCC,AABBCc。
c.最高能达到170厘米,最矮能达到160厘米。
14.a.若花器大小是由环境因素决定的,那么大×小将产生类似亲本的后代;若后代在某些方面区别于它们的亲代,那么就意味着花大小主要是由基因型决定的。
b.用大×小产生F2代,测出后代级别比例。如果是3:1或1:2:1就说明是由一对等位基因控制的;如果1:4:6:4:1就说明存在两对具有加性效应的等位基因。另外,统计F2代中极端类型的比例,亦可确定控制性状的基因对数。
15.a. A自交系为aabbcc;B自交系为AABBCC;
b. 180克;
c. 7个级别:120、140、160、180、200、220和240克;
d. 256株(平均64株中才有1株是AABBCC)。
16.
 ,4对基因的差别。
,4对基因的差别。
17.a.均受微效多基因控制的数量性状的遗传,且熟期性状主要受遗传基因控制,株高性状易受环境条件的作用。
b.由于熟期的狭义遗传力高,所以可推断,这10株熟期为120天的水稻,其基因型接近一致。
c.与b相反,由于秆高的狭义遗传力较小,所以尽管杆高都为80厘米,但遗传组成可能有较大的差异。
d.根据两性状遗传力的大小,可在早世代选熟期为120天的单株混合建系,到高世代再选株高接近80厘米的单株。
18.有效的性状遗传力大,而无效的遗传力小。
19.数量性状易受环境影响。
20.
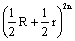 。
。
21.增加育种工作的预见性。凡遗传力高的性状,在杂种的早期世代选择效果显著;而遗传力较低的性状,在杂种晚期世代选择较好。
22.F2代植株平均高度:
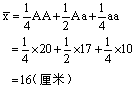
23.F2代果穗的平均长度:
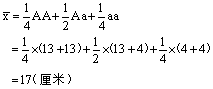
24.a.
 ;
;
b.
 ;
;
c.
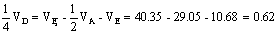 ;
;
d.
 。
。
25.a. 40厘米;
b.基因型AaBb、aaBB、AAbb的高度为40厘米;
c. F2代中40厘米高的基因型有
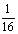 AAbb、
AAbb、
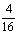 AaBb、
AaBb、
aaBB,共占
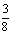 。
。
26.因为
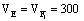 ,
,

所以
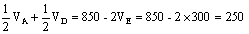 …………….①
…………….①
又因为
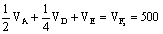
所以
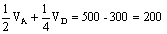 …………………………….②
…………………………….②
①-②得
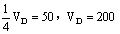
根据②得
 ,
,

27.

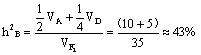

28.已知
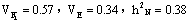
所以 a.
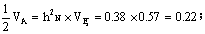
b.
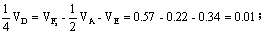
c.
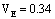
d.
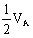 占总方差的百分率即狭义遗传力为38%,
占总方差的百分率即狭义遗传力为38%,
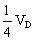 占总方差的百分率0.01/0.57=2%,VE占总方差的百分率0.34/0.57=60%。
占总方差的百分率0.01/0.57=2%,VE占总方差的百分率0.34/0.57=60%。
29.F1是四基因杂合体,故株高是:
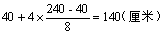 ,F2高度范围应是240厘米到40厘米之间。
,F2高度范围应是240厘米到40厘米之间。
30.极端类型的平均数为
 ,因此玉米籽粒的颜色可能由4对基因控制。
,因此玉米籽粒的颜色可能由4对基因控制。
二、判断对错
1.×;F1表现型方差越大,广义遗传力越小。
2.×;采用不同方法估算的遗传力,其值有一定的差异。
3.×;数量性状呈连续性变异,杂种后代不能明确地划组或分类。
4.√
5.√
6.×;遗传引起的变量占变异总量的百分比是遗传力。纯系的遗传力最小,等于0.
三、选择正确答案
1.b
2.①c;②d;③a;④b
3.①d;②c;③a;④b;⑤c
四、填空
1.简便易行;粗放致使估算的遗传力偏高
2.显著
3.较高
4.无显性偏差;存在部分显性作用;存在超显性作用
5.加性方差;显性方差;上位性方差;加性方差;显性方差;上位性方差;加性方差;表型总方差
6.11%;39%
7.
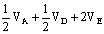
8.逐渐升高
9.0;环境方差;纯系;不同
五、名词解释
质量性状:生物体中,表现不连续变异的性状。
数量性状:生物体中,表现连续变异的性状。
微效多基因:指控制数量性状的基因,数量多,每个基因对表现型的影响较微,不能把它们个别的作用区别开来。简称微效基因。
主基因:对于性状的作用比较明显,控制着质量性状的基因。但在有些数量性状遗传中,主基因也在起作用。
修饰基因:是指一组效果微小的基因,能增强或削弱主基因对表现型的作用。
超亲遗传:在数量性状的遗传中,杂种第二代及以后的分离世代群体中,出现超越亲本性状的新表型的现象。
遗传率:生物群体中一个性状可遗传的变异占总变异量的百分率。
广义遗传率:遗传方差占总方差的比值。以百分数表示:
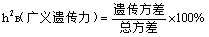
狭义遗传率:基因加性方差占表现型总方差的比值。计算公式:

表现型方差:杂种后代的表现型是由亲本的基因型和环境条件共同作用的结果,表现型方差是指两者共同作用引起的变异量。
基因型方差:是指基因型引起的变量,说明了基因型对表现型作用的能力。
加性方差:指等位基因间和非等位基因间的累加作用引起的变异量。
显性方差:指等位基因间相互作用引起的变异量。
上位性方差:指非等位基因间相互作用引起的变异量。
多基因假说:数量性状是许多彼此独立的基因作用的结果。各个等位基因的表现为不完全显性或无显性,或表现为增效和减效作用,各个基因对性状表现的效果微效,效应相等,作用是累加的,服从孟德尔的遗传规律。
环境方差:是指环境引起的生物体某性状的变异量。
表现型值:某性状表现型的数值。
基因型值:是指表现型值中的由基因型所决定的数值。
中亲值:指两亲本的中间值。
第三章
近亲繁殖和杂种优势
一、回答问题
1.雌雄配子来源于同一植株或同一花朵。
2.
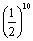
3.回交后代群体中基因型的纯合内容严格地受轮回亲本的控制,就是说纯合的内容完全为轮回亲本的基因型。而自交后代的基因型纯合却是多种多样的组合方式。在纯合进度上两者是相同的,都为
 。但就某一种基因型而言,自交的纯合率应为
。但就某一种基因型而言,自交的纯合率应为
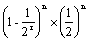 ,其纯合进度上明显小于回交。
,其纯合进度上明显小于回交。
4.这两种植物的花色都是受一对等位基因所控制。a的红花基因对白花基因为不完全显性,杂合时为粉红色。b的一对基因呈超显性遗传,杂合时呈红色。
5.相同处:杂交后代的某性状超过亲本。
不同处:杂种优势是在F1代性状超过亲本;基因型高度杂合时产生的优势;其优势不能稳定遗传。而超亲本遗传是在F2及以后世代中出现;基因型纯合是的超亲;能稳定遗传。
6.自交系间的杂交种比品种间的杂交种在F2代基因分离的更严重,因此杂种优势衰退的也就更严重。
7.a.杂交亲本的纯合性,只有两个基因型高度纯合的亲本的F1才能产生整齐一致的优势。
b.亲本杂交组合的选配,因为亲本不同,F1表现的优势也不相同。所以应选择配合力高的自交系做亲本。
c.杂交制种技术要简便易行,同时种子繁殖系数要高。
8.自花授粉植物由于长期自交,隐性性状得以表现。有害的性状已被自然选择和人工选择所淘汰。
9.由于杂种优势的大小与双亲基因型的纯合程度以及双亲的亲缘程度有关,在一定范围内双亲基因型的纯合程度愈大,亲缘关系愈远,杂种优势愈大。所以(A×D)×(B×C)产生的双交种杂种优势最强。
10.群体遗传组合纯合化;等位基因纯合化;各种性状稳定遗传。
11.由于种种因素的影响,一个纯系内的某些个体的遗传基础可能发生改变,导致有益性状的出现,因此可在多年推广的小麦品种中继续进行单株选择。
12.玉米属于异花授粉作物,自交F2等位基因纯合,一些隐性性状得以表现。而大田中这些隐性性状处于杂合状态而被遮盖,所以很少能够表现出白化苗、黄绿苗等隐性性状。
13.根据纯合率公式:

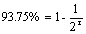 ;
;
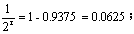
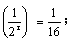 r=4,需连续自交4代。
r=4,需连续自交4代。
14.如果群体中杂交体的比例为100%,那么自交r代后杂合体的比例为
 。当群体中杂合体的比例为25%时,自交两代后杂合体的比例应为
。当群体中杂合体的比例为25%时,自交两代后杂合体的比例应为
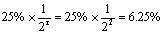 ,纯合体的比例为1-6.25%=93.75%。
,纯合体的比例为1-6.25%=93.75%。
15.根据公式:
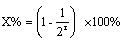 ,
,
,r=4,需回交4代。
16.如果进行自交利用公式
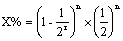 ,r=4;如果进行回交利用公式
,r=4;如果进行回交利用公式
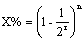 ,r≈1。
,r≈1。
17.两者相同均为
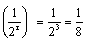
18.
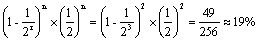 。
。
19.自交后代某一种基因型的纯合率公式为:
基因型Aabb和aaBb只存在一对基因的差异,所以n=1。
基因型AaBb中两对基因全呈杂合状态,所以n=2。
因此基因型Aabb、aaBb自交三代后aabb基因型的比例分别为
 ;
;
基因型AaBb自交三代后aabb基因型的比例则为
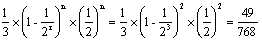 ;
;
群体中aabb基因型的总比例为

20.根据公式[1+(2r-1)]n=[1+25-1]3
=13+3×12×31+3×1×312+313
=1+93+2883+29791
=32768
三对基因全为杂合体的比例为1/32786;
三对基因中两对杂合一对纯合的比例为93/32786;
三对基因中群为纯合的比例为29791/32786。
二、判断对错
1.×;每自交一代,杂合体所占比例减少一半,逐渐接近于零,但总是存在的,不会完全消失。
2.×;一个纯系实际上是基因型纯合一致的群体。
3.×;花药植株是基因已经分离的单倍体植株。
4.×;自交对显隐性基因的纯合作用是一致的。
5.×;白化苗的出现是由于隐性等位基因纯合的结果。
6.×;杂种优势主要表现在数量性状上。
7.×;F1产生优势,F2衰退。
8.×;.应在一定范围内。
9.×;杂种优势的大小与环境条件也有着密切的关系。
10.×;主要采用自交系间的杂交种。
11.×;显性假说只注意到显性基因之的互补作用。
三、填空
1.
 ;
;

2.约翰逊
3.杂种优势来源于双亲基因型间的相互作用
4.近亲繁殖;自交(或自体受精);全同胞交配(或姊妹交);半同胞交配
5.杂交;品种间杂交;远缘杂交;品种间杂交;远缘杂交
6.显性假说;超显性;显性基因数目;异质等位基因对数
7.基因;优势;杂交;杂交种
8.群体中纯合体的比例增大,杂合体的比例减小;纯系;近交
9.基因对数;自交代数;选择的是否严格
10.纯合;隐性
11.各对基因是独立遗传的;各种基因型后代的繁殖能力相同
12.自交;回交;姊妹交
13.细胞核;父本
14.几种;一种
15.区分了遗传的变异和不遗传变异,提出选择遗传性变异的重要性。
16.F1超过双亲的平均值的百分数
17.大;双亲基因型的高度纯合
18.异质等位基因分离;基因型的纯合;相对纯合的遗传组成;纯系
四、名词解释
近亲繁殖:是指血统或亲缘关系相近的两个个体间的交配,也就是指基因型相同或相近的两个个体间的交配。
近亲系数:是指一个合子中两个等位基因来自双亲共同祖先的某一基因的概率。说明了近亲繁殖的程度。
回交:指杂种后代与两亲本之一的再次交配。
轮回亲本:指在回交中,被用来连续回交的亲本。
非轮回亲本:未被用来连续回交的亲本。
同质基因:指一对纯合的等位基因。
异质基因:指一对杂合的等位基因。
纯系:指从一个基因型纯合个体自交产生的后代。
纯系学说:是由约翰逊提出的。在自花授粉植物的天然混杂群体中,可分离出许多基因型纯合的纯系。在混杂群体中选择是有效的,纯系内选择是无效的。
杂种优势:指两个遗传组成不同的亲本杂交产生的杂种第一代,在生长势、生活力、繁殖力、抗逆性、产量和品质等方面比其双亲优越的现象。
显性假说:认为杂种优势是由于双亲的显性基因全部集在杂合体中所引起的互补作用。
超显性假说:认为基因型的异质结合可以增加基因间的作用,因而优于双亲的纯系结合。
永久性合体:为保持杂种优势,利用染色体结构变异,使杂种自交时,同质基因结合的合子会引起个体死亡,异质基因结合的合子可以生存,这种杂合体称永久杂合体。
第四章
基因突变
一、回答问题
1.自发突变——在没有人工特设的诱变条件下,由于外界环境条件的自然作用或由于生物体内的生理和生化变化而发生的突变。例如,1910年摩尔根在大量的野生型的红眼果蝇中发现了一只白眼突变的果蝇,这种由红眼突变为白眼就是自发突变。
诱发突变——生物在专门的物理和化学诱变因素或高温等其它因素影响下发生的突变。例如,水稻品种原丰早就是通过用
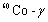 射线处理水稻,使其产生变异,再加以选择而育成的新品种。
射线处理水稻,使其产生变异,再加以选择而育成的新品种。
正突变——指显性基因突变为隐性基因。例如,水稻有芒基因A突变为无芒基因a。
反突变——指隐性基因突变为显性基因。例如,水稻无芒基因a突变为有芒基因A。
2.位于同—基因位点上各个等位基因称为复等位基因。根据人得A、B、O血型复等位基因的遗传知识,可以鉴别亲子间血缘关系。
3.在植物里,一个芽在发育的及早时期发生突变,这个芽长成枝条,上面着生的叶、花和果实跟其它枝条不同,这叫枝变或芽变。芽变在农业生产上占有重要位置。育种上每当发现性状优良的芽变就要及时扦插、压条、嫁接或用组织培养等方法加以繁殖,使它保留下来。不少果树新品种就是由芽变选育成功的。著名的温州早桔就是由温州蜜桔的芽变选育成的。
4.因为现存的生物都是经历长期自然选择进化而来的,它们的遗传物质及其控制下的代谢过程,都已达到相对平衡和协调。如果某一基因一旦发生突变,原有的协调关系不可避免地要遭到破坏或削弱,生物赖以正常生活的代谢关系就会被打乱,从而引起程度不同的有害后果,一般表现为生育反常。但是有些基因突变虽对生物体有害,却对人类有利,如雄性不育基因的突变。
5.亲缘关系相近的物种因遗传基础比较近似,往往发生相似的基因突变。说明当了解到一个物种或属内具有哪些变异类型,就能预见到近缘的其它物种或属内也同样存在相似的变异类型。突变的平行性对于研究物种间的亲缘关系、物种进化和人工的定向诱变都具有一定的意义。
6.体细胞中的隐性基因如果发生显性突变,当代个体就以嵌合体的形式表现出突变性状,要从其中选出纯合体,必须通过有性繁殖自交两代。如果发生隐性突变,虽然当代已成为杂合体,但突变性状因显性基因掩盖并不表现,要使其表现还须通过有性繁殖自交一代,性细胞发生的突变可以通过受精直接传递给后代,而体细胞则不能。突变了的体细胞在生长过程中往往受到抑制或最后消失,所以要保留体细胞的突变,需将它从母体上及时地分割下来加以无性繁殖。
突变性状的表现既因作物繁殖方式而不同,又因授粉方式而有别。当显性基因突变为隐性基因时,自花授粉植物只要通过自交繁殖,突变性状就会分离出来。异花授粉植物则不然,它会在群体中长期保持异质结合而不表现,只有进行人工自交或互交,纯合的突变体才有可能出现。
在自交的情况下,显性突变表现得早而纯合得慢,隐性突变与此相反,表现得晚而纯合得快。显性突变第一代表现,第二代纯合,而检出突变体在第三代。隐性突变第二代表现,第二代纯合,检出突变纯合体也在第二代。
现行突变的表现 隐性突变的表现
dd DD
↓ ↓
第一代(M1) Dd Dd
 ↓
↓
↓
第二代(M2) 1DD 2Dd 1dd 1DD 2Dd 1dd
 ↓
↓
↓
↓
第三代(M3) DD 1DD :2Dd :1dd dd
7.用具有隐性性状的纯种为母本,用由诱变处理的具有显性性状的纯种的花粉作父本进行杂交。
8.把矮杆连同原始亲本一起,种植在土壤和栽培条件基本均匀一致的条件下,如果变异体跟原始亲本大体表现相似,说明不是遗传的变异,反之,如果任然表现为矮杆,说明基因发生了变异。
突变体究竟是显性突变,还是隐性突变,这可以利用杂交实验加以鉴定。让突变体矮杆植株与原始亲本杂交,如果F1表现高秆,F2既有高秆,又有因分离而出现的矮杆植株,这说明矮杆突变是隐性突变,而不是显性突变。如属显性突变,也可用同样方法加以鉴定。
9.基因突变需要相当大的能量,因此细胞必须在得到大量能量以后,基因才可能突变。辐射就是很好的能量来源,电离辐射能使构成基因的化学物质直接发生电离作用。当电离辐射的射线碰撞基因任何分子时,射线的能量是基因任何分子的某些原子的外围电子脱离轨道,引起“原发电离”。在射线经过的道路上,在形成大量离子对的过程中所产生的电子,多数尚有较大能量,能引起“次级电离”。次级电离的结果,轻则造成基因分子结构的改组,产生突变了的新基因;重则造成染色体的断裂,引起染色体结构的畸变。所以在电离辐射的作用下,基因突变和染色体畸变往往是交替在一起的。
非电离辐射,紫外线的诱变作用在于被DNA吸收之后促使分子结构发生离析。紫外线的波长(3800—150
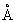 )比可见光略短,所以它的能量不足以使原子电离,只能产生激发作用。原子外围电子一旦活跃起来,就势必造成基因分子链的离析,这些分子链已经离析的基因在重新组合的时候,不免要发生差错,于是出现基因突变,紫外线还有间接诱变作用。比如用紫外线照射过的培养基去培养微生物,结果使微生物的突变率增加了。这是因为紫外线照射过的培养基内产生了过氧化氢(H2O2)。氢基酸经过氧化氢处理就有使微生物突变的作用。
)比可见光略短,所以它的能量不足以使原子电离,只能产生激发作用。原子外围电子一旦活跃起来,就势必造成基因分子链的离析,这些分子链已经离析的基因在重新组合的时候,不免要发生差错,于是出现基因突变,紫外线还有间接诱变作用。比如用紫外线照射过的培养基去培养微生物,结果使微生物的突变率增加了。这是因为紫外线照射过的培养基内产生了过氧化氢(H2O2)。氢基酸经过氧化氢处理就有使微生物突变的作用。
10.生化遗传研究指出,红色面包霉在合成生物所需物质时,要经过一系列的生化过程,而每一个生化过程又由一定的基因所控制。因此,只要综合分析大量的生化突变材料,就有可能阐明有关的基因功能及其作用的程序。现举例说明如下:
在红色包霉中,有一个突变型(a),给它提供精氨酸才能正常生长,否则就不能合成蛋白质。这说明它丧失了合成精氨酸的能力。另一个突变型(c)在有精氨酸的条件下能够正常生长,但不给精氨酸而只给瓜氨酸也能正常生长,这说明它能够用瓜氨酸合成精氨酸。第三个突变型(o)在有精氨酸或瓜氨酸的条件下能够正常生长,但不给这两种物质,而只给鸟氨酸也能生长,这说明它能够利用鸟氨酸最终合成精氨酸。
根据对上述几个突变型的测定,可以推论精氨酸的合成步骤与基因的关系大致为:
o c a
→鸟氨酸→瓜氨酸→精氨酸→蛋白质
由此可以看出,从鸟氨酸到精氨酸的合成至少需要A、C、O三个基因,其中任何一个基因发生突变,精氨酸就不能合成。
11.正常毛色:CC、CCh、CCa;喜马拉雅白化:ChCh、ChCa;白化:CaCa。
12. 父母的血型 孩子的血型
a. A A(IAi×IAi) O、A
b. AB O(IAIB×ii) A、B
c. AB A(IAIB×IAi) AB、A
d. A B(IAi×IBi) AB、A、B、O
只有a和b可能产生血型是O型的孩子。
13.母亲和孩子的血型是O型,那么血型为B的父亲的基因型一定为IBi,则第二个孩子为O型的机会为1/2。为B型的机会也为1/2。是AA型或AB型的机会均为0。
14.a. AY(黄): aa(黑)=2:1
b. AYA(黄): AYa(黄): Aa(鼠)=1:1:1,其中黄色:鼠色=2:1
c.AYa(黄): aa(黑)=1:1
d. AYA(黄): Aa(鼠)=1:1
e.AYA(黄): AYa(黄): Aa(鼠): aa(黑)=1:1:1:1
15.这是因为经过自然界长期自然选择保留下来的基因,对生物本身是有利的,在性状表现上大都是显性的。相反,多数突变是随机发生在基因内部的不同碱基座位上,使基因的功能形成不同的缺陷。这样,突变基因就不像正常型或野生型那样有适应能力,所以在性状表现上大多是隐性的。但突变等位基因的显隐性是相对的,有时也会出现显性突变基因,如在水稻的突变基因中,有半数左右是显性突变基因。
16.(1)h+→h-突变包括许多组氨酸基因,而h-→h+只涉及一个基因;
(2)h+→h-突变对于任何一个组氢酸基因来讲,包括许多突变点,而h-→h+只涉及一个位点。
17.X连锁突变在半合雄性个体上易于显现。
18. AABB 正常株 AABB
↓诱变处理 ↓诱变处理
AaBB AABb
↓
↓
 1AABB:2AaBB:1aaBB 1AAbb:2AABb:1AABB
1AABB:2AaBB:1aaBB 1AAbb:2AABb:1AABB
3正常株 :1突变株 × 1突变株 :3正常株
↓
F1 AaBb正常株
↓
 A_B_ AAbb aaBB aabb
A_B_ AAbb aaBB aabb
正常株 突变株
351 : 267
这主要是由于基因互作类型中的基因互补作用。
19.原因有二.(1)正突变往往是多方向性的,如A→a1,a2,……,而回复突变往往是a1或a2→A,而其余多数突变不一定回复变成A;(2)除了真正的严格回复突变外,还存在不少抑制突变。这一种拟突变型(假野生型)与野生型杂交后,后代还出现突变型。
20.a.不能结实; b. S1S3,S2S3; c. S1S3,S2S3,S1S4,S2S4
21.由于基因突变的多方向性产生的。
22.有两种可能性,即果实直感或芽变。前者可用种子繁殖选育新类型,后者可采用插条或嫁接育成新品种。
23.杂交变异;基因突变;染色体结构和数目的变异。
24.突变的频率低;突变的时期不定;突变的部位不等;突变的重演性;突变的可逆性;突变的多方向性;突变的有害性和有利性;突变的平行性。
25.基因重组引起的变异是原有基因的重新组合,基因本身并未发生改变。而基因突变是由于其本身的化学结构发生了改变。
26.普通燕麦是六倍体,野燕麦是四倍体。由于燕麦多倍体中不同组的染色体具有部分同源的关系,具有部分同源关系的染色体有少数基因作用是相同的,所以如果野燕麦和普通燕麦发生了基因突变,由于未突变基因的剂量效应,使突变的基因所控制的性状不能表现出来。因为矮燕麦是二倍体,不存在基因的剂量效应,所以矮燕麦的突变率要高于野燕麦和普通燕麦。
27.第一代:♀:♂=1:1;第二代:♀:♂=2:1。
二、判断对错
1.√
2.×,体细胞显性突变表现得早,分离出纯合突变体的时间晚。
3.3.√
4.4.√
5.√
6.×,在基本生长的菌株是野生型菌株。
7.√
8.√
9.√
10.√
三、填空
1.自花授粉不结实;同一;拮抗
2.化学性质;对性
3.甜粒;susu;非甜粒;SuSu
4.电离辐射;非电离辐射
5.伴性致死
6.完全培养基;基本培养基;基本培养基;精氨酸营养缺陷型
7.同一突变可以在同种生物的不同个体间多次发生;亲缘关系相近的物种因遗传基础比较近似,往往发生相似的基因突变
8.辐射剂量;辐射强度
9.不一样;高
10.嵌合体
11.Aa;aA
12.座位
13.可逆的
14.IAi;IBi;IAi
四、名词解释
基因突变:指染色体上某一基因位点内部发生了化学性质的变化,与原来的基因形成对性关系。例如,高杆基因D突变为矮杆基因d。因此,基因突变也称点突变。
突变体:由于基因突变而表现突变性状的细胞或个体称突变体,也称突变型。
正突变:即显性基因突变为隐性基因。
反突变:即隐性基因突变为显性基因。
复等位基因:位于同一基因位点上各个等位基因。
突变率:指突变体占观察个体数的比率。
芽变:在植物中,一个芽在发育的及早时期发生突变,这芽长成枝条,上面着生叶、花和果跟其他枝条不同,这叫枝变或芽变。
嵌合体:由不同基因型的细胞构成的生物体,可自发产生或人工产生。
正突变率:正突变发生的频率。
反突变率:反突变发生的频率。
自交不亲和性:指自花授粉不结实,而株间授粉却能结实的现象。
致死突变:导致个体死亡的突变。
隐性致死:指致死突变是隐性突变。
显性致死:指致死突变是显性突变。显性致死突变在杂合状态下即可死亡。
中性突变:有些基因仅仅控制一些次要性状,它们即使发生突变,也不会影响生物的正常生理活动,因而仍保持其正常的生活力和繁殖力,为自然选择保留下来。这类突变一般称为中性突变,例如:水稻芒的有无。
M1:由诱发突变当代长成的植株。
M2:用M1繁殖的后代为M2。
营养缺陷型:它与正常野生型即原养型的区别在于丧失了合成其中某种生活物质的能力。当一但提供这种生活物质,它同野生型一样又能正常生长。
原养型:指正常的野生型。
自发诱变:在自然情况下产生的突变。
诱发突变:由人们有意识地应用一些物理、化学因素诱发的突变。
完全培养基:在微生物学中,补充有酵母菌浸出物、蛋白胨等营养物质的基本培养基。可以用来培养各种营养缺陷型。
伴性致死:指致死突变发生在染色体上。
大突变:指突变效应表现明显,容易识别,这类突变一般发生在控制质量性状的基因上。
微突变:指突变效应表现微小,较难观察,一般发生在控制数量性状的基因上。
第五章染色体结构的变异
一、回答问题
1.染色体结构变异是DNA分子上较大的区段发生了变化;基因突变是指染色体上某一基因位点内部发生了化学性质的变化,即DNA分子上某一区段内的一个或几个核苷酸发生了变化。如果染色体结构变异非常微小,两者就难以区分。
2.根据杂合体减数分裂时偶线期或粗线期染色体联会的形式:
缺失:有瘤或环状的突起,染色粒、染色节的缺少。
重复:有瘤或环状的突起,染色粒、染色节的增多。
倒位:出现倒位圈,或染色体中间配对,两端不配对。
异位:十字形图像、四体环、四体链等。
3.孢母细胞减数分裂期间,后期Ⅰ会产生染色体桥和断片。后期Ⅱ断片消失,桥断裂成两个缺失的染色体。最后形成的四分孢子中,两个败育,两个可育。
4.有两种可能,一是父本植株在配子形成过程中,少量的G基因突变为g;二是发生了含有G基因染色体片段的缺失。证明的方法有以下几种:
a.回交验证:将F1代中的绿株与亲本的紫株纯合体(GG)回交,再将回交一代(BC1)严格自花授粉。如果是染色体缺失,自交后代将呈现紫株与绿株之比为6:1;若属基因突变,自交后代呈3紫株:1绿株的比例。图示如下:
染色体缺失情况下:(绿株个体只有一个g基因)go×GG
↓
BC1 Gg Go
↓
↓
3G_:1gg 3G_:1oo(死亡)
基因突变情况如下:gg×GG
↓
BC1 Gg
↓
3G_:1gg
以上结果是有条件的,即假定缺失不影响雄配子的育性。
b.花粉检查:如果属染色体缺失,那么杂交F1代中表现隐性的植株,一定是缺失杂合体。由于雄配子一般对缺失更为敏感,所以花粉中可能有部分败育。
c.细胞学鉴定:观察F1隐性植株花粉母细胞减数分裂粗线期(或双线期)染色体构象,若出现缺失圈(中间缺失)或非姊妹染色单体不等长(顶端缺失)的二价体,就可断定为染色体缺失。
5.因为倒位杂合体的大多数含倒位交换染色单体的配子是不育的,它所能产生的交换配子数显著地减少,连锁基因重组率就降低。
6.原因1:染色体发生顶端缺失后,两条姊妹染色单体在断点处融合,或两条具有断头的染色体在断点处融合,都可形成具有双着丝点的染色体。在细胞分裂后期这些染色体被拉向两极时,会有“桥”的现象出现。
原因2:染色体发生臂内倒位后,倒位杂合体在倒位区段内非姊妹染色单体间发生了一次交换,从而形成双着丝点的缺失染色体,后期Ⅰ也会有“桥”的现象出现。
7.倒位杂合体表现为部分不育,而异位杂合体为半不育。
8.





9.不一定。这是一个倒位杂合体,如果这一对染色体的非姊妹染色单体间未发生交换,或者在倒位区段内发生了一次单交换,那么只能产生两种可育的配子,即A·BCDEF和a·bedcf。如果交换发生在倒位区段之外,或在倒位区段内发生了双交换,那么产生的多种配子都是可育的。
10.产生4种可育配子,即ABCDEF、ABCdEF、abeDcf.abedcf。图解如下:


粗线期 后期Ⅰ 四分孢子
11.
 如果减数分裂后期Ⅰ联会在一起的四条染色体呈相邻式分离可产生四种配子,即ABCDEF FEDONM、ABCPQR MNOPQR、ABCDEF ABCPQR和FEDONM MNOPQR。四种配子中都含有缺失染色体,因此都是不育的。如果这四条染色体呈交替式分离,可产生两种类型配子,即ABCDEF MNOPQR和ABCPQR MNODEF。这两种配子都不缺少正常染色体的任何区段,因而都能发育成正常可育的配子。(如图)
如果减数分裂后期Ⅰ联会在一起的四条染色体呈相邻式分离可产生四种配子,即ABCDEF FEDONM、ABCPQR MNOPQR、ABCDEF ABCPQR和FEDONM MNOPQR。四种配子中都含有缺失染色体,因此都是不育的。如果这四条染色体呈交替式分离,可产生两种类型配子,即ABCDEF MNOPQR和ABCPQR MNODEF。这两种配子都不缺少正常染色体的任何区段,因而都能发育成正常可育的配子。(如图)
12.
 甲植株:1 3 4 2
甲植株:1 3 4 2
1 2 4 3
乙植株:1 4 3 2
1 2 3 4
13.
 a.倒位
a.倒位
b.
1 2 3 7 8
1 2 3 7 8
14.有臂间倒位和臂内倒位两种。两者的杂合体在减数分裂前期Ⅰ都能形成倒位圈,当非姊妹染色单体在倒位圈内发生交换时,臂内倒位杂合体可能产生双着丝点染色单体,随着出现“后期桥”的构象。臂间倒位则无此现象。
15.由于倒位杂合体可使倒位区段内连锁基因的重组率下降。所以倒位发生在包含i基因的区段。因为该基因与两相邻基因的交换值为y-i,4.5%和i-s,1.2%,远远小于它的正常值35%和9%。
16.2n=20本应联合成10个二倍体,9个联会体可能有一个是易位形成的四价体。后期Ⅰ的染色体桥可能是有一对染色体是臂内倒位杂合体,并且在倒位区段内发生了单交换形成双着丝点染色体而产生的。
17.
 a.
a.
b.产生两种可育配子:ABCD EFGH和ABGH DCFE。
18.两条正常染色体和两条易位染色体在减数分裂的偶线期联会成“十”字形象,终变期交叉端化成四体环或四体链。后期Ⅰ这四条染色体如果呈交替式分离,产生的配子全可育。如果呈相邻式分离,产生的配子全不育。两种分离方式的几率大致相等,所以表现为半不育。
19.
AA BB CC C C A A
B B
B B
 CB CA AB AA CC
CB CA AB AA CC
20.
 甲: 乙:
甲: 乙:
↓ ↓
可育 abcd abgh abcd abfe
 配子 efgh dcfe efgh dcgh
配子 efgh dcfe efgh dcgh
abcd abgh abcd abgh
F1 efgh dcfe efgh dcfe
abcd abcd abfe abfe
efgh efgh dcgh dcgh
 联会
联会
 a a e e
a a e e
b b f f
c c g g
d d h h
正常的二价体联会与十字形联会的比例为1:3.
21.
 a.
a.
d D
a b c Y X W
×
A B C y x w
Zz
 b.①A B c dw x y z②a b C ZD Y X W③A B C ZD Y X W
b.①A B c dw x y z②a b C ZD Y X W③A B C ZD Y X W
④a b c d w x y z
22.产生4种可育配子,遗传组成分别为:
a. 1-2-·-3-4-5-6-7-8 a -b -·-c-d g -h -·-m-n
b. 1-2-·-3-4-5-6-7-8 A-B-·-M-N G-H-·-C-D
c. 1-2-·-3-6-5-4-7-8 a - b-·-c-d g - h-·-m-n
d. 1-2-·-3-6-5-4-7-8 A-B-·-M-N G-H-·-C-D
23.a. G·E//MF M·N//G·N
b.相邻式分离,形成的配子染色体组成为:
(G·F M·F)(G·N M·N) (G·F G·N) (M·N M·F)均不能存活。
c.交替式分离形成的配子组成为:
(G·F M·N)(G·N M·F)都能成活。
24.无叶舌、黄绿苗、正常可育:有叶舌、绿苗、半不育=1:1.
25.a.由于在减数分裂的花粉母细胞中观察到了十字形结构,因此可断定这是两对非同源染色体相互易位的结果。
b.该分裂相是减数分裂前期I的偶线期或粗线期。
c.植株花粉母细胞中的六个四合体是六对未发生变异的染色体的组成,而十字形结构是由两对相互异位染色体组成,因此2n=6Ⅱ+1Ⅳ=16,体细胞中的染色体数为16。
d.易位杂合体的遗传效应之一是半不育现象,即所形成的配子中有半数是可育的,半数是不育的。所以该植株的花粉只有半数可育。
26.

甲 A B C D
A B C D
乙 A C B D
A C B D
丙 A D C B
A D C B
27.利用染色体缺失产生的假显性现象进行基因定位。如果染色体上某一显性基因染色体片段缺失而丧失作用,其隐性的等位基因得以表现。进行细胞学检查缺失发生哪条染色体的哪一部位,就可确定该基因的位置。
28.表示玉米第5染色体和第9染色体的一个相互易位。
29.产生含有重复缺失染色体的无活力配子,导致倒位杂合体的部分不育。
30.

31.a.两亲本的全部配子都携带完整的基因组。
b.纯合亲本的全部配子和杂合亲本的一般配子具有完整的基因组,总计占全部产生配子的75%。
32.
 设原始物种染色体区段顺序为ABCDEFGHIJ,而ABCHGFEDIJ则为原始物种发生的一次倒位所形成的变种:
设原始物种染色体区段顺序为ABCDEFGHIJ,而ABCHGFEDIJ则为原始物种发生的一次倒位所形成的变种:
 A B C I J
A B C I J
 而ABCHGFIDEJ是在述变种的基础上又发生一次倒位所形成的另一个新变种:
而ABCHGFIDEJ是在述变种的基础上又发生一次倒位所形成的另一个新变种:
A B C H G F J
33.可能是中间缺失造成的。F1基因型+e,如归一条同源染色体上“+”所在区段缺失,就使果蝇表现型变为e,即黑檀体型。在这种果蝇唾腺染色体上找到缺失环可以证实。
34.AA亲本为易位纯合体,F1是易位杂合体。该异位点不在载有A-a基因的染色体上。因此 亲本AA的遗传基础为AATT,F1为AaTt。
35.两亲本之一为易位杂合体,另一个为正常体。F1为易位杂合体,同时也是A-a基因杂合体,易位点不在基因A-a所载的染色体上,遗传组成为AaTt。
36.第9染色体缺失的断头点与C基因的距离为10个遗传单位,在♀cc×♂缺失杂合体Cc的杂交中,缺失杂合体在减数分裂时,C基因与缺失断头区段内可能发生非姊妹染色单体交换,这样载有显性基因的C的染色体就成为正常染色体,其所形成的小孢子成为可育,能参与正常的受精活动。图示如下:

♀[正常的cc纯合体] ×♂[缺失的杂合体Cc]

 c c C c
c c C c
缺失杂合体在减数分裂时同源染色体的非姊妹染色单体进行交换,交换值为10%,故可产生如下几种配子:
C C c c
未交换型 交换型 交换型 未交换型
45%5% 5% 45%
败育可育 败育 可育
在可育的花粉中,C型配子占可育配子10%。所以受精后就有10%的籽粒的基因型为Cc,表现有色。
37. YYTT × yytt
↓
YyTt × yytt
↓ ↓
a.形成的可育配子:YT yt Yt yT yt
b.测交子代:基因型 表现型 比例
YyTt 黄胚乳半不育 40%
yytt 白胚乳完全可育 40%
Yytt 黄胚乳完全可育 10%
yyTt 白胚乳 半不育 10%
38. TTAA×ttaa
↓
TtAa × ttaa
↓
TtAa Ttaa ttAa ttaa
半不育的A 半不育的a 正常的A 正常的a
1528 290 272 1454
重组率=(290+272)/(1528+290+272+1454) ×100%=15.8%。基因A/a距离易位点15.8个遗传单位。
39. FfBbTt ×ffbbtt
↓
无条纹、正常色脉、半不育 FBT 96
有条纹、棕色脉、完全可育 fbt 99
无条纹、正常色、完全可育 FBt 9
有条纹、棕色脉、半不育 fbT 12
有条纹、正色脉、半不育 fBT 63
无条纹、棕色脉、完全可育 Fbt 67
有条纹、正常色脉、完全可育fBt 3
无条纹、棕色脉、半不育 FbT 2
从测交结果可知,F和B基因连锁与一条染色体上,而且易位也发生在该染色体上。
双交换值=(3+2)/351=0.014
f-b单交换值:(63+67)/351+0.014=0.384
b-t单交换值:(9+12)/351+0.014=0.074
 f、b、t三者关系为 _f 38.4 b 7.4 t_
f、b、t三者关系为 _f 38.4 b 7.4 t_
40.由于倒位杂合体的倒位区段内连锁基因的重组率下降,因此可知培养物1和培养物2中的果蝇都是X射线引起的倒位结果。与正常重组率相比,又可知培养物1倒位发生在m和f区段,培养物2的倒位发生在w和m区段。
41.易位发生在含有A基因的染色体上,而且与A基因相聚(15%+15%=30%)30个遗传单位。
二、判断对错
1.√
2.×;不一定,如果缺失的区段特小,含有缺失染色体的花粉也可能全部可育。
3.√
4.×;倒位是染色体片段的正常直线顺序的颠倒,染色体片段并未转移。
5.×;缺失杂合体和臂内倒位杂合体都只能产生含缺失染色体的不育配子。
6.√
7.×;只有在倒位区段内发生单交换的臂内倒位杂合体,后期I才可能形成染色体桥,臂间倒位杂合体不形成染色体桥。
8.×;倒位杂合体的大多数重组型配子是不育的,所以重组型个体数减少。
9.√
10.×;臂内倒位能形成桥,臂僭倒位则不能。
11.√
12.×;叫转移
13.×;非同源染色体间染色体片段的转移
14.×;相互易位杂合体相邻式分离后,配子中的染色体为非同源染色体间的重复-缺失类型,与倒位杂合体形成的同源染色体间的重复-缺失类型不同。
15.×;25%
16.√
17.×;辐射强度对染色体的两次断折有一定的影响。
18.√
19.×;把含有标记基因的染色体节段易位到W染色体上,后代中凡是含有标记性状的个体一定会发育成雌性,因为雌性的性染色体为ZW型。
20.×;这是玉米第2染色体和第4染色体的一个易位,故应产生8个正常的二价体和一个“四体环”。
三、选择正确答案
1.c
2.d
四、填空
1.数目;位置
2.染色体缺失造成的假显性现象;隐性突变
3.倒位圈;染色体桥
4.染色体易位
5.正常个体;易位杂合体;易位纯合体;1:2:1;易位杂合体
6.“十”字形;相邻式;交替式
7.抑制交换的倒位区段;该倒位区段内的一个隐性致死基因;该倒位区段外的一个16区A段的重复
8.同源染色体;顺接重复;反接重复;剂量效应;位置效应;剂量;剂量效应;剂量;位置;位置效应
9.缺失;重复;倒位
10.辐射剂量;辐射剂量的平方
五、名词解释
先断后接假说:在自然条件下,或人为的用某些物理因素或化学药剂处理细胞,染色体会发生折断。折断后染色体可能按原来的直线顺序再次接合起来,也可能发生错接,后者就会造成染色体结构的变异。这在细胞学中称先断后接假说。
缺失:是指染色体的某一区段丢失了。
顶端缺失:指染色体缺失的区段是某臂的外端。
中间缺失:指染色体缺失的区段是某臂的内段。
双着丝点染色体:指具有两个着丝点的染色体或染色单体。
染色体断片:指无着丝点的染色体片段。
缺失杂合体:某个体的体细胞内杂合有正常染色体及其缺失染色体。
缺失纯合体:指某个体的缺失染色体是成对的。
染色体缺失环:在缺失杂合体的减数分裂过程中所观察到的染色体联会现象。由于正常染色体与缺失染色体配对时,有一部分区段不能和缺失部分配对,从而形成一个向一边鼓起的小环。
假显性:在显隐性杂合体中,由于载有显性基因的染色体发生了缺失,显性基因随着缺失片段丢失了,从而使隐性基因控制的性状得以表现。
重复:指染色体多了自己的某一区段。
顺接重复:是指某区段按照自己在染色体上的正常直线顺序重复了。
反接重复:是指某区段在重复时颠倒了自己在染色体上的正常直线顺序。
剂量效应:细胞内某基因出现的次数越多,表现型效应就越显著。
位置效应:一个基因随着染色体畸变而改变了它对邻近基因的位置关系,从而改变它的表型效应的现象。
染色体重复环:在重复杂合子减数分裂过程中,重复染色体和正常染色体联会时,由于重复区段不能和正常染色体配对,从而形成一个环或瘤。
倒位:是指染色体的某一区段的正常直线顺序的颠倒。
臂内倒位:倒位区段在染色体的某一臂范围内,也称一侧到位。
臂间倒位:倒位区段涉及染色体的两个臂。也称两侧倒位。
染色体桥:双着丝点染色体在减数分裂后期当两着丝点分别移向两级时,染色体横跨赤道板的图像。
染色体倒位圈:在倒位杂合子减数分裂过程中,正常染色体和倒位染色体的倒位区段相对应的一段旋转180形成一个圈,再和倒位染色体进行联会。
易位:指染色体的一个区段移接到另一条非同源染色体上。
简单易位:指某染色体的一个臂内区段,嵌入非同源染色体的一个臂内的现象。也称转移。
相互易位:两个非同源染色体都折断了,而且这两个折断了的染色体及其断片随后又交换地重新结合起来。
半不育:花粉和胚囊各有50%是败育的。
易位杂合体:某个体的体细胞内杂合有正常染色体及其易位染色体。
易位纯合体:指个体的易位染色体是成对的。
“十”字形联会:在相互易位杂合体中,四条染色体具有部分同源性,但其中没有任何两条染色体完全相同,在减数分裂前期染色体配对时,四条染色体联会成一个“十”字。
染色体融合:两条相互易位的染色体中,一条得到正常染色体的很小一段,而另一条却得到正常染色体的绝大部分。这样的一个易位杂合体在产生配子时,很容易丢失小染色体,它的自交子代群体内,就有可能出现少了一对染色体的易位纯合体。
相邻式分离:易位杂合体减数分裂时,四条染色体配对构型中,相邻的两条染色体被拉向同一级。
交替式分离:在易位杂合体四个染色配对的构型中,想间隔的两个染色体抵达同一级。
半合状态:一对等位基因只存在其中一个的情况,如那些坐落在性染色体上的基因或由于缺失造成的结果。
体细胞联会:体细胞中同源染色体联会现象。
不等交换:发生在联会时略有参差的两个染色体区段之间的交换,结果使一个染色单体具有重复染色体区段,而另一个则缺失了部分区段。
重复一缺失染色体:指一条染色体重复了自己的某一区段的同时,另一区段发生了缺失。
双杂合体:具有杂合基因位点的某一染色体畸变杂合体。
ClB染色体:果蝇的一种特殊的X染色体,其上含有一个倒位区段,倒位区段内有一隐性致死基因,倒位区段之外有一个16区A段的重复区段。
ClB测定:利用ClB//X+雌蝇测定X染色体上基因的隐性突变频率的方法。
第六章 染色体数目的变异
一、回答问题
1.同一染色体组中的各条染色体的形态、结构和连锁群都彼此不同,但它们却构成了一个完整而协调的体系,缺少其中任何一个都会造成不育或性状变异。这就是染色体组的最基本特征。
2.a. 2n-1=41;b. n=21;c. 2n+2=44;d. 2n-2=40;e. 2n+1+1=44
3.n=102/2=51;x=102/6=17
4.远缘杂交的不孕性是指亲缘关系较远的物种杂交不结实或不受孕的现象。而远缘杂种的不实性是指远缘物种杂交所产生的杂种不能再繁殖后代的现象。
5.根据同源多倍体某些器官、组织或细胞具有趋大性的特点加以判别。如一般情况下同源多倍体多的叶片保卫细胞比正常二倍体大,但单位面积内的、气孔数则相对较少。
6.a. 19条;b.利用染色体加倍技术;38条
7.
染色体数 目的变异 |
染色体组(X) 及其染色体 |
合子染色体数(2n)及其组成 |
主要联 会方式 |
染色体 组数 |
染色体 组类别 |
染色体 |
|
整 倍 体 |
二倍体 |
A=a1a2a3 |
2x |
AA |
a1a1a2a2a3a3 |
3Ⅱ |
B=b1b2b3 |
2x |
BB |
b1b1b2b2b3b3 |
3Ⅱ |
E=e1e2e3 |
2x |
EE |
e1e1e2e2e3e3 |
3Ⅱ |
同 源 |
三倍体 |
A=a1a2a3 |
3x |
AAA |
a1a1a1a2a2a2a3a3a3 |
3Ⅲ |
四倍体 |
同上 |
4x |
AAAA |
a1a1a1a1a2a2a2a2a3a3a3a3 |
3Ⅳ |
异 源 |
四倍体 |
A=a1a2a3 B=b1b2b3 |
4x |
AABB |
(a1a1a2a2a3a3) (b1b1b2b2b3b3) |
6Ⅱ |
六倍体 |
A=a1a2a3 B=b1b2b3 E=e1e2e3 |
6x |
AABBEE |
(a1a1a2a2a3a3) (b1b1b2b2b3b3) (e1e1e2e2e3e3) |
9Ⅱ |
三倍体 |
同上 |
3x |
ABE |
(a1a2a3)( b1b2b3)( e1e2e3) |
9Ⅰ |
非 整 倍 体 |
亚倍体 |
单体 |
A=a1a2a3 B=b1b2b3 |
2n-1 |
AAB(B-1b3) |
(a1a1a2a2a3a3) (b1b1b2b2b3) |
5Ⅱ+Ⅰ |
缺体 |
同上 |
2n-2 |
AA(B-1b3) (B-1b3) |
(a1a1a2a2a3a3) (b1b1b2b2) |
5Ⅱ |
双单体 |
同上 |
2n-1-1 |
AA(B-1b2-1b3) |
(a1a1a2a2a3a3) (b1b1b2b3) |
4Ⅱ+Ⅰ+Ⅰ |
超倍体 |
三体 |
A=a1a2a3 |
2n+1 |
A(A+1a3) |
a1a1a2a2a3a3a3 |
2Ⅱ+Ⅲ |
四体 |
同上 |
2n+2 |
A(A+2a3) |
a1a1a2a2a3a3a3a3 |
2Ⅱ+Ⅳ |
双三体 |
同上 |
2n+1+1 |
A(A+1a2+1a3) |
a1a1a2a2a2a3a3a3 |
Ⅱ+Ⅲ+Ⅲ |
8.杂种细胞内一个染色体组来自糖槭,一个来自羽叶槭。减数分裂时,两者难以正常配对,从而导致染色体分离的紊乱,不能产生可育的配子,使杂种不育。如果将杂种细胞染色体加倍,使之变为异源四倍体,可成为可育的植株。
9.在偶倍数异源多倍体的细胞内,由于每种染色体组都有两个,同源染色体都是成对存在的,减数分裂时能像二倍体一样联会成二价体。所以可表现与二倍体相同的遗传规律。
10.a.加倍后,可得到完全可育的纯合体;b.基因的显隐性都能得以表现;c.可用于研究非同源染色体间的部分同源关系。
11.在同源三倍体的细胞内,每个同源组含三条染色体。在减数分裂时,这三条染色体或联会成三价体或一个二价体和一个单价体。因此,后期I的分离主要是2/1的不均衡分离。结果造成同源三倍体的配子染色体组合成分的不平衡从而表现高度不育。
12.不对。因为除了同源四倍体(复式)外,复式三体(AAa)按染色体随机分离后形成的配子比例为1 AA : 2 Aa :2A:1a,与隐性亲本回交后,显隐性性状之比为5[A] :1 [a]。
13.孤雌生殖、孤雄生殖和假受精现象等可以产生单倍体种子,从而形成单倍体植株。单倍体植株在细胞减数分裂时染色体分离紊乱,不能形成可育配子,因而不能繁殖后代。
14.因为隐性性状纯合的几率二倍体大于同源四倍体。
15. 黑麦(RR)×小麦(AABBDD)
↓
ABDR
↓染色体加倍
小黑麦(AABBDDRR=28Ⅱ)×小麦(AABBDD=21Ⅱ)
2n=7X=AABBDDR=49=21ⅡT+7IR
↓
AABBDD+IR×AABBDD-I(2n-1)
↓(小麦单体)
2n-1+1=20Ⅱ+IT+IR
↓
↓
↓
选择出 2n=20Ⅱ+ⅡR=21Ⅱ
(其中:T代表小麦染色体、R代表黑麦染色体)
16.杂种F1具有A物种的5条染色体和B物种的7条染色体,共12条染色体。可能杂种F1产生了少量为减数的花粉粒,含12条染色体。这些花粉粒与B物种卵细胞的7条染色体(n=7)结合,产生了含19条染色体的植株,其染色体组成为(5 Ia+7 IIbb)。这种植株可产生了极少数含12条染色体的花粉和胚囊(5 Ia+7 Ib)。自交后产生24条染色体的新植株(2n=5IIaa+7IIbb)。
17.秋水仙素可阻碍分生细胞内纺锤丝的形成,使复制后的染色体不能被拉向两极,结果不能形成两个子核,导致细胞内染色体数目加倍。
18.母本为单式三体,父本为复式三体。控制红花基因R对白花基因r为显性。该基因距离着丝点很近,减数分裂时服从染色体随机分离。单式三体(Rrr)的母本形成的胚子的种类和比例为2Rr:1R:2r:1rr。复式三体(RRr)的父本形成的花粉基因型的种类和比例为1RR:2Rr:2R:1r。母本的全部配子都能参与受精。而父本中只有n型配子参与受精,自交后将有(2Rr:1R:2r:1rr)(2R:1r)=5[R]:1[r]。即红花:白花=49:10≈5:1。
19.小麦2n=AABBDD×黑麦2n=RR
↓
杂种ABDR
↓染色体加倍
AABBDDRR小黑麦
20.不对。利用单体进行隐性基因定位时,应以隐性性状的双体植株(2n)分别与各个显性纯合的单体(2n-1)杂交。杂交后代中,单体植株的单体染色体只能来自双体亲本。
21.整倍体:①单倍体
②多倍体:a.同源多倍体
b.异源多倍体 偶倍数异源多倍体
奇倍数异源多倍体
c.同源异源多倍体
非整倍体:①超倍体a.三体;b.双三体;c.四体
②亚倍体a.单体;b.双单体;c.缺体
22.将隐性aa的植株分别与小麦的21个显性单体杂交:
a.a基因在单体染色体上:
P A表现型单体×a表现型双体
[(n-1)II+IA]↓ [(n-1)II+IIaa]
F1[(n-1)II+IIAa] [(n-1)II+Ia]
A表现型双体 a表现型单体
杂交F1中双体植株全为A表现型,单体植株全为a表现型。
b. a基因不在单体染色体上:
P A表现型单体×a表现型单体
[(n-1)IIAA+I]↓[(n-1)IIaa+II]
F1 [(n-1)IIAa+II] [(n-1)IIAa+I]
A表现型 A表现型
杂交F1中无论是双体植株还是单体植株都是表现为A表现型,没有a表现型出现。
23.该物种属同源四倍体;每个同源组中有4四条同源染色体,每个染色体组包含12条染色体;2n=12×4=48。
24.a.同源染色体;b.纯合体;c.复等位基因;d.DNA分子;e.中间着丝点染色体;f.核型;g.多线染色体;h.秋水仙素。
25. 一粒小麦 拟思卑尔脱山羊草
×
AA(2n=14) BB(2n=14)
↓
杂种AB
↓染色体加倍
二粒小麦AABB×方穗山羊草DD(2n=14)
↓
杂种ABD
↓染色体加倍
斯卑尔脱小麦(2n=AABBDD)
↓
↓基因突变
↓
普通小麦(2n=AABBDD)
26.a.萝卜单倍体染色体应为9。
b.萝卜2n=2X=BB=9Ⅱ×甘蓝2n=2X=RR=9
↓
2n=2X=BR=18I
↓少量未减数的配子结合
萝卜甘蓝2n=4X=BBRR=36=18Ⅱ
27.前者联会成14个二价体,7个单价体(14Ⅱ+7Ⅰ)。后者联会成7个二价体,21个单价体(7Ⅱ+21Ⅰ)。
28.普通小麦有两个A,两个B和两个D染色体组,现已知道这三组染色体从1A、1B和1D到7A、7B和7D是部分同源的。既是部分同源,就有一部分基因相同,所以普通小麦某一单位性状常常由3对独立基因控制。辐射诱变时,往往以二倍体突变频率较高。这是因为基因突变一般是独立发生的。在异源六倍体的三对基因中,如果其中某一个基因发生隐性突变(显性→隐性),突变后的基因会受其它五个不曾突变的基因的掩盖。如果基因发生显性突变(隐性→显性),也常常由于剂量效应的影响,而不能表现出来。二倍体只有一对基因,其中一个发生突变,就比较容易分离表达出来。异源四倍体界于二倍体和异源六倍体之间。
29.a.若所缺的那条染色体属S染色体组:
[(SSTT)-1]×SS
↓
[(SS-1)+T]=11Ⅱss+Ⅰs+12ⅠT=11Ⅱ+13Ⅰ
若所缺的那条染色体属T染色体组:
[(SSTT-1)]×SS
↓
[SS+(T-1)]=12ⅡSS+11ⅠT
所以所缺的那条染色体属S染色体组。
b.12个二价体和11个单价体(12IIⅡSS+11ⅠT)。
30.玉米是二倍体生物,其配子内只有一个染色体组。当其中的一条染色体丢失后,染色体组的完整性遭到破坏,形成的n-1型的配子一般不能正常发育,所以它的子代群体内也就不会出现亚倍体品系。普通小麦是异源六倍体,他的配子中含有三个染色体组。产生的n-1型配子内虽然缺失了某一个染色体组的某一条染色体,但所缺失的那条染色体的功能可由另一染色体组的相应染色体所补充,所以n-1型配子还能正常发育。其后代可以出现亚倍体系列。
31.两对互补基因控制花色,其中一个亲本为单体,丢失的染色体上载有显性基因,即Abb×aaBB。
32.a.双体(48条)、红花:单体(47条)、白花为1:1。
b.花色全为红色。理论上单体与双体比值相等。
33.a.复式,染色体随即分离;b.单式,染色体随机分离;c.三式,染色单体随机分离。
34.A为复式三体,B为单式三体;孢母细胞均按染色体随即分离;n+1型花粉不参与受精,参与受精的n+1型胚囊与n型胚囊之比为1:3。
35.三体亲本的基因型为AAa,双体亲本为Aa,基因按染色体随机分离,三体的n+1卵子只要1/4参与受精,即(n+1):n=1:3。
36.a.复式三体;服从染色体随机分离;所有的配子均可以正常可育。
b.单式三体;服从染色体随机分离;所有的配子均正常可育。
37.a.复式三体(AAa);服从染色体随机分离;参与受精的n+1型雌配子与n型雌配子之比为1:3。
b.单式三体(Aaa);服从染色体随即分离;参与受精的n+1型雌配子与n型雌配子之比为1:3。
38.两种非整倍体均为单体;红花非整倍体的基因型为A,白花非整倍体的基因型为a,红花双体的基因型为Aa。
39.第一种情况,花色基因不在该缺体染色体上。第二种情况,花色基因在该缺体染色体同源组上。
40.先天愚型患者产生的配子中,一半为只含一条21号染色体的正常配子,另一半配子中含有两条21号染色体。假如对方为正常者,受精后,孩子中有一半的可能为先天愚型患者。
41.a.发生在精子;b.难以确定;c.发生在卵子。
42.AABBD;35条;
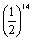 。
。
43.a. AA:Aa=15:12:1;AA:Aa:aa=1:12:15。
b. AA:Aa=1:1;Aa:aa=1:1。
44.A/a基因所在染色体和三体Ⅲ有关。
45.如果该基因不在某三体染色体上,应有下列杂交结果:
FF×ff
↓
Ff×ff
↓
1 Ff : 1 ff
即测交后抗病与感病之比为1:1。对照题中结果,只有与三体-10杂交后的测交子代中两种性状之比不等。因此,可以认为该基因位于第10条染色体上。
46.检查减数分裂的细胞。如果是同源四倍体,则联会复合体包含4条染色体即四价体;如果是异源四倍体,则仍联会成二价体。
47.如果基因Yb1/ yb1(或者Yb2/ yb2)涉及某单体染色体(M-U)应有下列遗传过程:
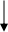

 P:
P:
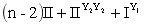 ×
×
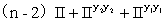


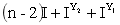
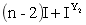
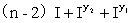
F1:
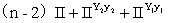
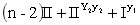
绿色双倍 绿色单倍
测交:
×


Ft:

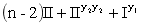
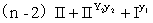
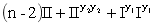
绿株 白肋株
可见,若该基因涉及某单体染色体,那么经上述杂交后,即测交子代中绿株与白肋株之比为1:1。对照题给数据推测,该基因可能位于O染色体上。
48.复式三体AAa染色体随机分离后形成的配子种类和比例为1AA:2Aa:2A: 1a。单式三体Aaa的配子为2Aa:1aa:1A:2a.
Aaa×AAa
↓
2AAAa:5AAaa:2Aaaa:1AAA:8AAa:8Aaa:1aaa:2AA:5Aa:2aa
49.a. 1红色:1无色; b. 1红色:2无色;c. 2红色:1无色
50.Bbb→(2Bb:1B:1b:1bb)2→3[B]:1[b]
51.由题意可知,该基因按染色体随机分离
a.基因型++ra形成的雌配子的种类和比例为1++:2+ra:2+:1ra,能够参与受精雌配子的种类和比例为1++:2+ra:6+:3ra。因此♀++ra×♂rara→1++ra:2+rara:6+ra:3rara,后代中隐性表现型所占的比例为3/12。
b. ++ra形成的雌配子的种类和比例分别为1++:2+ra:2+:1ra,能够参与受精的雌配子的种类和比例为1++:2+ra:6+:3ra;能够参与受精的雄配子的种类和比例为2+:1ra,因此♀++ra×♂rara→11[+-]:1[rara].后代中隐性表现型所占的比例为1/12。
c.基因型+rara所形成的雌配子的种类和比例为2+ra:1+:2ra:1rara.能够参与受精的雌配子的种类和比例为2+ra:3+:6ra:1rara.因此♀+rara×♂rara→2+rara:3+ra:6rara:1rarara。后代中隐性表现型所占比例为7/12。
52.如果基因a在该三体染色体上,aa x AAA杂交F1代中的三体植株基因型为AAa,回交后代显隐性比例应为5[A_]:1[aa]。
53.当两组基因均为染色体随机分离时:
 35[B]→1225[A][B]
35[B]→1225[A][B]
35[A]
1 [b]→ 35 [A][b]
35[b]→ 35[a][B]
1[a]
1 [b]→ 1 [a][b]
当两组基因均为染色单体随机分离时:
187[B]→34969[A][B]
187[A]
9 [b]→1683 [A][b]
187[B]→1683[a][B]
9 [a]
9 [b]→81 [a][b]
当两组基因中,一组为染色体随机分离,一组为染色单体随机分离时:
187[B]→6545[A][B]
35[A]
9 [b]→315[A][b]
187[B]→187[a][b]
1 [a]
9 [b]→ 9 [a][b]
54.四体占5%,三体占50%,双体占45%。
55.a.假设c基因在第6染色体上,那么当F1代群体中的三体植株与隐性亲本回交应有:
CCc×cc→(1CC:2Cc:2C:1c)(c)→1CCc:2Ccc:2Cc:1cc
即正常叶形与马铃薯叶形之比为5:1;2n+1与2n之比为1:1.
b.若c基因不在第6染色体上,应有:Cc×cc→1Cc:1cc
即两种表现型之比为1:1.
56.如果su基因在地10染色体上,F1代中三体的基因型应为Sususu。自交后代应出现单式三体的表现型分离比例,与实验结果不符。如果su基因不在第10染色体上,F1代所有个体的基因型都为Susu。自交后淀粉质与甜质之比为3:1。与题给的结果(1758:586≈3:1)相符。所以Su/su基因不在第10染色体上。
57.共同8种表现型,其比例分别为:
[Py][Lg][Ji]353/363 [Py][Lg][ji] 352/363
[Py][lg][Ji]352/363 [py][Lg][Ji] 352/363
[Py][lg][ji]35/363 [py][Lg][ji] 35/363
[py][lg][Ji] 35/363 [py][lg][Ji]1/363
58.
 ;
;
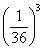 ;
;
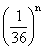
59.

60.a.后代中二倍体的比例1/2。
b.后代中基因型为Bbb的三体植株比例为1/3。
二、判断对错
1.√
2.×;代表双单体。
3.√
4.×;相反。
5.×;不一定。对双倍体物种来说,单倍体就是一倍体,而多倍体则不然。
6.×;不一定。如同源四倍体的单倍体细胞内染色体是成对存在的。
7.×;小麦是异源六倍体。
8.×;比正常二倍体染色体数少几条或多几条的个体叫非整倍体。
9.×;该基因表现为染色单体随机分离。
10.√
11.√
12.×;应为
 。
。
13.×;分离不出亚倍体系列。
14.√
15.×;异源四倍体的花粉植株一般是不结实的。
16.×;该三体是单式三体,该基因距离着丝点较近。
17.×;单式三体;供试基因距着丝点较近,按染色体随机分离;所有的配子全部能参与受精作用。
18.×;由于普通小麦的许多基因具有异位同效作用,所以不能直接利用两点或三点测验进行基因定位。
19.×;一定是异源多倍体。
20.√
三、选择正确答案
1.d
2.c
3.①c; ②d; ③b; ④c
4.b、c、d
四、填空
1.单体;三体;双单体;单体;三体
2.偶倍数异源多倍体;两个
3.两种;染色体随机;染色单体随机
4.4;AAA、AAa、Aaa、aaa
5.15AA:12Aa:1aa
6.
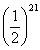 ;
;
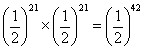
7.不同的偶倍数异源多倍体
8.二倍体;同源三倍体;异源三倍体;异源六倍体;异源五倍体;异源七倍体。
9.整倍体变异;非整倍体变异;染色体结构变异
10.染色体随机;1Aa:1aa;染色单体随机;1AA:12Aa:15aa
11.单倍体;高度不育;表现;纯合的;纯系
12.普通;双二倍
13.21;21;三体
14.同源三倍体;瓜;染色体加倍;同源四倍体;二倍体父本
15.生物学的方法;物理学的方法;秋水仙素
五、名词解释
染色体组:在二倍体生物的配子中所包含的形态、结构和功能上彼此不同的一组染色体。是多倍体物种染色体的组成成分。
一倍体:具有一个染色体组的生物体。
二倍体:每一个细胞中,具有两个染色体组的生物体。
三倍体:具有三个染色体组的生物体。
多倍体:具有三个或三个以上染色体组的生物体。
单倍体:具有配子染色体数(n)的个体。
同源多倍体:染色体组来源于同一物种的多倍体。
异源多倍体:染色体组来源于不同物种的多倍体。
偶倍数异源多倍体:每种染色体组都有两个,同源染色体都是成对的。
奇倍数异源多倍体:指不同的偶倍数多倍体种间杂交的子代。
双二倍体:即异源四倍体。对于两个基因组都呈二倍体状态的生物。
倍半二倍体:指细胞内一部分染色体组是双倍的,还有一个染色体组却是单倍的。
同源联会:在异源多倍体中,同一染色体组内的同源染色体发生联会。
异源联会:在异源多倍体中,不同染色体组内的染色体由于存在部分同源关系而发生联会的现象。
节段异源多倍体:指某些异源多倍体的不同染色体组间虽然也是部分同源的,但同源部分的程度很高。
同源-异源多倍体:是使异源多倍体的染色体数在加倍而形成的多倍体。
染色体同源组:指在同源多倍体中,几个形态、大小、结构相同的染色体。
染色体部分同源组:指在异源多倍体中,来自不同染色体组的几个染色体存在着部分同源的关系,称这些染色体为一个部分同源组。
提早解离:在多价体中每两个染色体之间只是局部联会的,交叉较少,联会松弛,就可能过早地松懈为更低价体。
不联会:每个同源组的染色体出现了单价体。即,没有发生联会。
染色体随机分离:当基因在某一同源组的染色体上距离着丝点较近,以致基因与着丝点之间很难发生非姊妹染色单体的交换时,该基因随染色体的分离而分离的现象。
染色单体随机分离:当基因在某一同源染色体上距离着丝点较远,以致基因与着丝点之间容易发生非姊妹染色单体的交换时,该基因随染色单体分离而分离的现象。
整倍体:染色体数是染色体基数的整倍数的个体。
非整倍体:比正常合子染色体数(2n)多或少一跳以至若干条染色体的细胞或个体。
超倍体:在正常合子染色体数(2n)基础上,增加一条以至若干条染色体。
亚倍体:在正常合子染色体数(2n)基础上,减少一条以至若干条染色体。
三体:在正常合子染色体数(2n)基础上,某对染色体外加一条染色体。
双三体:在正常合子染色体数(2n)基础上,某两对染色体都外加一条染色体。
单体:在正常合子染色体数(2n)基础上,缺少了一条染色体。
双单体:在正常合子染色体数(2n)基础上,某两对染色体同时少了一条染色体。
四体:在正常合子染色体数(2n)基础上,某对染色体外加两条染色体。
缺体:在正常合子染色体数(2n)基础上,某对染色体全都丢失了。
双体:指具有正常合子染色体数(2n)的个体。
双倍体:指具有合子染色体数(2n)的异源多倍体。
单价体:在减数分裂中期没有配对的单个染色体。
三价体:在减数分裂中,三个染色体的联会。
多价体:三价体以及三价体以上的称多价体。
异位同效基因:异源多倍体不同染色体组之间常存在部分同源的关系,因此,在不同的染色体组的染色体上存在着控制相同性状的基因。
远缘杂交不孕性:生物进行远缘杂交时,由于不同种属的个体亲缘关系太远,配子间不相适应,以致不能正常受精、胚胎不能正常发育的现象。
远缘杂交不实性:亲缘关系较远的动物或植物彼此杂交所得的杂种,因其生理功能不协调,生殖系统遭受扰乱而不能繁殖后代或繁殖能力很弱的现象。
第七章细菌和病毒的遗传
一、回答问题
1.a.细菌和病毒繁殖世代所需要的时间短,每个世代以分钟或小时来计算。
b.一支试管可以储存数以百万计的细菌和病毒,易于管理。细菌代谢旺盛,繁殖快,可在短时间内积累大量产物,为基因作用的研究和对基因进行化学分析提供了条件。
c.细菌和病毒的遗传物质比较简单,更适宜作基因结构和功能以及基因工程的研究。
d.细菌和病毒属于单倍体,所有突变都能立即表现出来,这样便于研究基因的突变。由于细菌繁殖快,短时间能形成大量的个体,所以即使基因突变频率低,也能检出突变体。
e.细菌可以生活在基本培养基上,易于获得营养缺陷型,也易于测知各种营养缺陷型所需要的物质,是研究基因作用的好材料。
f.细菌和病毒的结构简单,教易对其分析研究,可用作研究高等生物的简单模型。
2.大肠杆菌含有一个染色体,呈环状,是裸露的DNA分子。玉米的一个染色体组中含有10条染色体,其染色体中含有DNA、RNA、蛋白质和少量的拟脂与无机物质。
3.转化:是指某些细菌通过其细胞膜摄取周围供体的DNA片段,并将此外源DNA通过重组参入到自己染色体组中的过程。
接合:是指两菌体细胞直接接触,遗传物质从供体(雄性)转移到受体(雌性)的过程。
转导:是指以噬菌体为媒介所进进行的细菌遗传物质重组的过程。
性导:是指接合时由F'因子所携带的外源DNA整合到细菌染色体的过程。
共同点是转化、接合、性导和转导都是细菌获取外源遗传物质的途径;转导与转化、性导、接合的主要不同之处在于转导是以噬菌体为媒介所进行的细菌遗传物质的重组过程。
4.(1)先将两种不同营养缺陷型菌株培养液分别放入戴维斯U型管的两臂,由于两种菌株被滤片所隔开,排除了接合重组的可能性,如仍有原养型菌株出现则考虑以下原因。
(2)分析混合的培养液中可能的过滤性因子。以DNA酶处理混合培养液,如无原养型菌株出现,可以确定是由转化所致。
(3)如果以DNA酶处理混合培养液后,仍有原养型出现,而且菌液中有噬菌体存在,则可能是转导的结果。
(4)若菌液中无噬菌体出现,则可以进一步推测是否属于基因突变。
5.这个位点距离染色体的转移原点应该远。因为只有离原点远,这样这个位点前的Hfr基因可先进入F-,然后可用选择性培养基消除Hfr供体。
6.
7.a.因为大肠杆菌的重组是在受体染色体与供体染色体片段间发生的,不产生交互重组子,因此选择的一定是受体细胞。根据题给基因型(假定顺序已知,并且距离较远)和被选择的多数重组子是a-b+c+d-,可以推知,重组方式为:



可见,a+b+c-d+为供体菌。
b.从以上可以看出,在特定标记基因的重组中,其它非选择标记基因总是受体基因型的频率高。依此可以判定大肠杆菌的性别(供体和受体)
8.这个大肠杆菌是溶源性的。因为λ噬菌体是温和性噬菌体,它侵入大肠杆菌后,大肠杆菌并不裂解。
9.
 a.三个基因顺序是: leu ara ilvH
a.三个基因顺序是: leu ara ilvH
b.这个转导片段的最大极限是从ilvH位点到leu和ara位点之间。
10.选择性地杀死供体菌,保留下来的都是接合后代。
11.a-b之间的重组率=
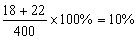
b-c之间的重组率=
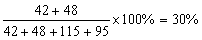
a-c之间的重组率=
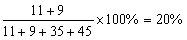
因此三基因的顺序及距离为: b←10→a← 20→c
12.a.真核生物DNA中G+C比例变动大,而原核生物DNA中G+C的比例较为稳定。
b.真核生物每个细胞中DNA的含量大,而原核生物每个细胞中DNA含量小。
c.真核生物DNA含有大量的重复顺序,而原核生物DNA仅有很少的重复顺序或没有重复顺序。
d.真核生物DNA基因中有较多的插入顺序,及内含子,故由它转录的mRNA要经过加工,切除内含子,才能成为有功能的mRNA。而原核生物DNA中没有插入顺序,其mRNA一般不需要加工,而且转录和翻译是在同一地点和几乎在同一时间进行的。
13.a. F因子以游离状态存在; b. F因子整合到细菌染色体上。
14.用不同的Hfr菌株进行中断杂交试验。
15.重组率(a-b)=

重组率(b-c)=
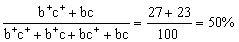
重组率(a-c)=

 三基因的顺序及图距: b←31→ a←27→c
三基因的顺序及图距: b←31→ a←27→c
16.交换值(m-r)=(520+474+162+172)/10342=12.8%
交换值(m-t)=(853+965+520+474)/10342=27.2%
交换值(t-r)=(853+965+162+172)/10342=20.8%
所以三基因之间的距离和顺序为: m←12.8→r←20.8→t
17.P22侵染细胞后,细菌染色体断裂成片段,在形成噬菌体颗粒时,偶尔(千分之一的机率)错误地把细菌染色体片段包装在噬菌体蛋白质外壳内,其中并不包含噬菌体的遗传物质。这种假噬菌体称为转导颗粒。因为决定感染细菌能力的是噬菌体的外壳蛋白质,所以这种转导颗粒可以吸附到细菌上。这种颗粒既然不携带噬菌体基因,对受体细菌就没有有害的影响。当转导颗粒将它的内含物注入受体细菌后,形成一个部分二倍体,导入的基因经过重组,整合到宿主的染色体上。
18.烈性噬菌体侵染细菌后可使细菌裂解,温和噬菌体侵染细菌后,并不使细菌裂解。
19.不能。因为噬菌体只能在有代谢活动的细菌中增殖。不能利用乳糖作为碳源的细菌是不能在以乳糖作为唯一碳源的培养基上存活的。
20.Hfr菌株的基因是按一定的线性顺序依次进入F-菌株的。也就是说,染色体从原点开始是以直线方式进入F-细胞的。基因位点离原点越近,进入F-细胞越早,反之距离原点越远的基因进入F-菌株的机会也越少。
21.高等生物通过亲本产生的雌雄配子的结合而将遗传物质传给下一代。而细菌则是通过两菌体细胞直接接触,将遗传物质从供体细胞转入受体细胞,或者通过菌体细胞吸收外来DNA分子片段;或通过某些噬菌体来传递遗传物质;还可以由F´因子将其所携带的外源DNA整合到细菌的染色体上。
22.重组率(ala-pro)=(2100+840+1400+840)/14840=0.35
重组率(pro-arg)=(2100+420+840+840)/14840=0.28
重组率(ala-arg)=(840+1400+420+840)/14840=0.24
a.ala-pro、pro-arg、ala-arg间的图距为别为35、28、24。
b.由于细菌染色体为环形的,因此,推断这三个基因的相对位置是;
 ala 35 pro
ala 35 pro
24 28
23.只有通过两种菌株的直接接触才能实现遗传物质的转移,即接合的结果。
24.
 a.
a.
b.“1”菌株与F-杂交,可选W基因作标记;
“2”菌株与F-杂交,可选X基因作标记;
“3”菌株与F-杂交,可选P基因作标记;
“4”菌株与F-杂交,可选F基因作标记;
“5”菌株与F-杂交,可选S基因作标记;
25.因为Hfr×F-出现的重组个体数最多,F+×F-出现重组体数少,而Hfr×Hfr、Hfr×F+、F-×F-都不会出现重组体。所以:a为F+;b为F-;c为F-;d为Hfr;e为Hfr;f为F-。
26.e—c—a—d—b—str
27.
28.欲想在这个杂交后代中选出重组体,应将所有的杂交子代培养在含有链霉素的培养基中,以便杀死所有的Hfr菌株。在含有链霉素的基本培养基中能够生存的菌株必是亮氨酸原养型Leu+,同时也是链霉素抗性strr。所以,只要在这种培养基中选出leu+菌株也一定就是strr菌株。因此应以leu+基因为选择标记。
29.基因v与t或p较近,而与b和m较远。
30.d—a—e—c—b
31.细菌的染色体为环状,单数交换使环状染色体打开,产生一个线性染色体,这种细胞是不能成活的。只能发生偶数交换才能产生环状染色体的重组体,而含有这样环状染色体的细胞是能存活的。
32.F+×F-→全部为F+; Hfr×F-→但大部分仍为F-,极少数为Hfr。
33.P4X: lac—gal—his—arg—xyl—ilu—thr
Ra-2: ilu—xyl—arg—his—gal—lac—thr
KL198:arg—xyl—ilu—thr—lac—gal—his
为起点 为终点
34.这是根据五个Hfr菌株与各自的营养缺陷型所做的中断杂交试验结果绘制的细菌染色体图。H菌株的转移起点为thr,终点为thi。1号菌株的转移起点为thr,终点为pro。2号菌株的起点为pro,终点为lac。3号菌株的起点为pur,终点为gal。312菌株起点为thi,终点为gly。
35.三个位点的顺序为cba;bc较近,ba离得远。转导片段的大小在a到b与c之间。
36.前种菌株是Hfr,细菌内附加体即F因子整合到染色体上,F因子与细菌染色体同步进行;后者是F+菌株,其内的F因子的存在是独立自主状态。不依赖宿主染色体而独立进行分裂。
37.供体DNA片段的大小、形态、浓度和受体细胞的生理状态:
a.转化片段大小要适当,如肺炎双球菌的成功转化的外源DNA片段至少需要有800个碱基对。枯草杆菌的转化片段最少需要16000个碱基对。
b.外源DNA片段存在的形式必须是双链的。
c.需要有一定浓度的外源DNA片段。
d.被转化的受体细胞必须处于感受态。
二、判断对错
1.√
2.×;F´因子是指携带有大肠杆菌染色体上某些基因的F因子。
3.×;不超过10个。
4.×;使原来的F-成为F+,原来的F+仍是F+。
5.×;P22可以转导沙门氏菌染色体的任何部分。
6.√
7.√
8.√
9.√
10.√
11.√
12.×;用F-表示。
13.√
14.×;发生偶数交换才能产生遗传的重组体。
15.×;细菌不裂解。
16.√
三、选择正确答案
1.d
2.a
3.d
4.a
5.b
四、填空
1.极高的基因转移率;极高的自然整合率
2.转化;接合;性导;转导
3.转化DNA片段;受体细胞
4.超数感染
5.裂解
6.F因子;供体菌;F+
7.F+;F-;高频重组菌株;Hfr
8.二倍;部分二倍体或部分合子
9.环状
10.供体外;受体内
11.接合;F´;染色体
12.一条;单倍
13.Hfr;同步
五、名词解释
F-菌株:不具有F因子的菌株。
F+菌株:具有一个自主状态的F因子的菌株。
Hfr菌株:细菌染色体组上整合有F因子的菌株。
F因子:细菌的性因子。是决定细菌性别的附加体。
F´因子:携带有细菌染色体的一些基因的F因子。
烈性噬菌体:能引起寄主细胞裂解的噬菌体。
温和性噬菌体:侵入寄主细胞后,不使寄主细胞裂解的噬菌体。
部分二倍体:当F+或Hfr的染色体进入F-后,在一个短时间内,F-细胞中对某些位点来说,总有一段二倍体的DNA。这样的细菌称为部分二倍体。
供体外基因子:在一个部分二倍体的细菌细胞中所包含的来自供体的部分染色体。
受体内基因子:一个部分二倍体细胞中的原有染色体的一部分,和供体外基因子同源。
菌落:在固体培养基上由单个微生物细胞长成的细胞集团或菌丝体。
噬菌斑:在含细菌的固体培养基上,噬菌体使细菌细胞裂解而形成的空斑。一般认为每个噬菌斑都是由一个噬菌体颗粒一再感染增值裂解发展而成的。
整合:非寄主染色体DNA(例如质粒、温和噬菌体、转化因子)并入寄主染色体上的过程。
转化:是指某些细菌(或其它生物)能通过其细胞膜摄取周围供体的染色体片段,并将此外源DNA片段通过重组渗入到自己染色体的过程。
接合:在原核生物中,接合是指两菌体细胞直接接触后,遗传物质从一个菌体细胞中转移到另一个菌体细胞中去的过程。
性导:是指接合时由F´因子所携带的外源DNA整合到细菌染色体的过程。
转导:是指以噬菌体为媒介所进行的细菌遗传物质重组的过程。
并发性导:对几个紧密连锁的基因同时由一个F´因子转移到受体细胞中的过程。
细菌裂解:当烈性噬菌体的遗传物质经中空尾部进入宿主细胞,遂即破坏宿主细胞原有的遗传物质,并转而合成大量的噬菌体遗传物质和蛋白质,组装成许多新的子噬菌体,最后使菌体细胞破裂,释放出数百个子噬菌体。
溶源性:噬菌体的遗传物质与寄主细菌的遗传物质相结合并协同复制的现象。
原噬菌体:温和性噬菌体感染细菌后,噬菌体整合于寄主染色体中,并且作为染色体的一部分而协同复制,这种噬菌体称原噬菌体。
超数感染:一个细菌受一个以上噬菌体感染的现象。
普遍性转导:对细菌染色体的任何不同部位都能转导的现象。
质粒:原指一切染色体外的遗传结构,但现在习惯上专指存在于细菌等微生物细胞中的染色体以外的遗传结构。
附加体:在细胞中或是以游离状态,或是以与染色体相结合的状态存在着的遗传结构。它们的存在给细胞带来一定的遗传性状,但它们的缺失并不照成细胞的死亡。
中断杂交试验:一种用来研究细菌接合过程中基因转移方式的试验方法。把接合中的细菌在不同时间取样,并把样品猛烈搅拌以分散接合中的细菌,然后分析受体细菌的基因顺序。是大肠杆菌等细菌中用来测定基因位置的一种方法。
寄主或宿主:被寄生物所感染或寄生的生物。
回复突变:突变基因转变成为野生型基因的过程。
假噬菌体:由寄主细胞的DNA和噬菌体的外壳所组成的颗粒。
影印培养法:使在一系列培养皿的相同位置上出现相同菌落的接种培养方法。把长有细菌菌落的培养皿倒过来印到绒布上,接着把无菌培养皿倒过来,在绒布上印一下,将每一菌落接种到相应的位置上。
第八章遗传物质的分子基础
一、回答问题
1.从化学上分析,真核生物的染色体是核酸和蛋白质的复合物。其中核酸主要是脱氧核糖核酸(DNA),在染色体上平均约占27%,其次是核糖核酸(RNA),约占6%;蛋白质占66%,是由组蛋白与非组蛋白构成的,两者的含量大致相同,但根据细胞的类型与代谢活动,非组蛋白的含量与性质变化较大。此外还有少量的拟脂与无机物质。
2.分子遗传学以拥有大量直接和间接证据,说明DNA是主要的遗传物质。
(1)DNA作为主要遗传物质的间接证据。
①每个物种不同组织的细胞不论其大小和功能如何,它们的DNA含量是恒定的。而且配子中的DNA含量正好是体细胞的一半,多倍体系列的一些物种,其细胞中DNA的含量随染色体倍数的增加也呈现倍数性的递增。
②DNA在代谢上是比较稳定的。
③DNA是所有生物的染色体所共有的,从噬菌体,病毒,直到人类的染色体中都含有DNA。
④用不同波长的紫外线诱发各种生物突变时,其最有效波长均为2600
 人,这与DNA所吸收的紫外线光谱是一致的。
人,这与DNA所吸收的紫外线光谱是一致的。
(2)DNA作为主要遗传物质的直接直接。
①细菌的转化:1928年,格里费斯(Griffith,F.)首次将一种类型的肺炎双球菌RII转化为另一种类型RIII,实现了细菌遗传性状的定向转化。16年后,阿委瑞(Avery,O.T.)等用生物化学方法证明这种转化物质是DNA。
②噬菌体的侵染与繁殖;赫尔歇等用同位素32P和35S分别标记T2噬菌体的DNA和蛋白质。然后用标记的T2噬菌体分别感染大肠杆菌,经10分钟后,用搅拌器甩掉附着于细胞外地噬菌体外壳。发现用32P标记,放射性活动见于细菌内而不被甩掉并可传递给子代。用35S标记,放射性活动大部分见于被甩掉的外壳中,细菌内只有较低的放射性活动,但不能传递给子代。这样看来主要是由于DNA进入细胞内才产生完整的噬菌体。所以说DNA是具有连续性的遗传物质。
3.①有两条互补的多核苷酸链,彼此以一定的空间距离,在同一轴上相互盘旋起来,很像一个扭曲起来的梯子。
②在DNA双链中,一条链的走向从5′到3′,另一条链的走向从3′到5′。两条链呈反向平行。
③A与T以两个氢键配对相连,G与C是以三个氢键配对相连。
④各对碱基上下之间的距离为3.4
,每个螺旋的距离34
,也就是说,每个螺旋包括10对碱基。
4.a.这条链是DNA。
b.如以之为模板,形成互补DNA链,它的碱基顺序为:T-G-G-C-A-A-A-T
c.如以之为模板,形成互补RNA链,它的碱基顺序为:U-G-G-C-A-A-A-U
5.T的含量为0.26。因为总得含量为1,G的含量和C的含量相同,C的含量为0.24。G与C的总含量为0.48,则A和T的含量为1-0.48=0.52,因为A和T含量相等,所以T的含量为0.26。
6.不能;因为DNA上碱基的排列顺序与碱基比例无直接关系。
7.相同点:都是多核苷酸的多聚体。
不同点:a.DNA含脱氧核糖,RNA含核糖;b.DNA中含A、T、C、G四种碱基,RNA含A、U、G、C;c.DNA为双链,RNA多为单链。
8.DNA复制,首先是从它的一端沿氢键逐渐断开,当双螺旋的一端已拆开为两条单链时,另一端仍保持双链状态,复制时按着5′到3′方向,RNA聚合酶以DNA为模板,合成一小段的含几十个核苷酸的引物RNA,然后DNA聚合酶才能开始起作用,按5′到3′端的方向合成DNA片段,也就是引物RNA的3′端与DNA片段的5′端接在一起。然后DNA聚合酶1将引物RNA除去,并且弥补上DNA片段,最后由DNA连接酶将DNA片段连成一条连续的DNA链。各自形成一条新的互补链与原来的模板单链互相盘旋在一起,恢复了DNA的双链结构。这样随着DNA双螺旋的完全拆开,就逐渐形成了两个新的DNA分子与原来的完全一样,DNA的这种复制方式称为半保留复制。
大多数RNA病毒是单链的。这种RNA的复制一般是以自己为模板合成一条互补的单链,通常称病毒原有的起模板作用的RNA分子链为“+”链,而将新复制的RNA分子链为“-”链,这样就形成了双螺旋的复制类型。然后这个“-”链又从“+”链模板释放出来,它也以自己为模板复制出一条与自己互补的“+”链,于是形成了一条新生的病毒RNA。
9.mRNA的第一个功能是把DNA上的遗传信息准确无误的转录下来。另一个功能是负责将它携带的遗传信息在多核糖体上翻译成蛋白质。TRNA在蛋白质合成过程中,将氨基酸搬运到核糖体上,并根据mRNA的遗传密码依次准确地将它携带的氨基酸联结成多肽链。rRNA一般与核糖体蛋白质结合在一起,形成核糖体。如果把rRNA从核糖体上除掉,就会发生塌陷。核糖体是合成蛋白质的场所。
10.转录成的mRNA是:5′AUGUUACCGGGAAAAUAG3′。反密码子是:UACAAUGGCCCUUUUAUC。肽链含有的氨基酸:甲硫氨酸、亮氨酸、脯氨酸、甘氨酸、赖氨酸。
11.用3H尿核苷标记大变形虫(Amoeba proteus)的核DNA转录的mRNA,然后把标记了的mRNA移植到未标记的变形虫中,用放射自显影术追踪标记的分布。结果发现,大部分标记了的mRNA随后转移到细胞质中,也就是说DNA先把遗传信息转录给RNA,然后在翻译成蛋白质。这个实验证明蛋白质不是直接从DNA合成的。
12.因为不缺U、A、G和C,所以仍可继续合成RNA和蛋白质。
13.蛋白质的合成过程,也就是遗传信息从DNA→mRNA→蛋白质的转录和翻译的过程,以及遗传信息从DNA→DNA的复制过程,这就是分子生物学的中心法则。中心法则所阐述的是基因的两个基本属性:自我复制和蛋白质合成。关于这两个属性的分子水平的分析,对于深入理解遗传及变异的实质具有重要的意义。
进一步的研究发现,在许多RNA的肿瘤病毒中,存在有反转录酶,它可以用RNA为模板,合成DNA。这一发现增加了中心法则中遗传信息的原有流向,丰富了中心法则的内容。另外还发现大部分RNA病毒可以把RNA直接复制成RNA。由于这些新的发现,可以把遗传信息的传递方向增添新的流向。
14.每一个氨基酸都由连续的三个核苷酸所决定的,这三个核苷酸叫做三联体密码。碱基与氨基酸两者之间的密码关系,显然不可能是一个碱基决定一个氨基酸。因此,一个碱基的密码子是不能成立的。如果是两个碱基决定一个氨基酸,那么两个碱基的密码子可能的组合将是42=16种。这比现存的20种氨基酸还差4种,因此不敷应用。如果是每三个碱基决定一种氨基酸,这三个碱基的密码子可能的组合将是43=64种。这比20种氨基酸多出44种。所以产生这种过剩的密码子,可以认为是由于每个特定的氨基酸是由一个或一个以上的三联体密码所决定的。从1961年开始,经过大量的试验,分别利用64个已知三联体密码,找出了它们对应的氨基酸。1966-1967年,全部完成了这套遗传密码字典。
15.一个氨基酸由一个以上的三联体密码所决定的现象,称为简并。简并现象对生物遗传的稳定性具有重要的意义。同义的密码子越多,生物遗传的稳定性越大。因为一旦DNA分子上的碱基发生突变时,突变后所形成的三联体密码,可能与原来的三联体密码翻译成同样的氨基酸,因而在多肽链上就不会表现任何变异。
16.①经典遗传学认为基因是一种化学实体,是成直线排列在染色体上。按照经典遗传学对基因的概念,基因具有下列共性:
1)基因具有染色体的主要特性:自我复制与相对稳定性,在有丝分裂和减数分裂中有规律的进行分配。
2)基因在染色体占有一定位置(位点),并且是交换的最小单位,即在重组时不能再分割的单位。
3)基因是以一个整体进行突变的,故它又是一个突变单位。
4)基因是一个功能单位,它控制着正在发育有机体的某一个或某些性状,如红花、白花等。可以把重组单位和突变单位统称为结构单位。这样,基因既是一个结构单位,又是一个功能单位。
②分子遗传学认为,基因在DNA分子上,一个基因相当于DNA分子上的一定区段,它携带有特殊的遗传信息,这类遗传信息或者被转录为RNA(包括mRNA、tRNA、rRNA);或者被翻译成多肽链(指mRNA);或者对其他基因的活动起调控作用(调节基因、启动基因、操纵基因等)。在一个基因的区域内,仍然可以划分出若干个起作用的小单位。按照现代分子遗传学的概念,重组、突变、功能这三个单位应该分别是:
1)突变子,它是性状突变时,产生突变的最小单位。一个突变子可以小到只是一个核苷酸对。
2)重组子,在发生性状的重组时,可交换的最小单位称为重组子(交换子)。一个交换子只包含一对核苷酸。
3)顺反子(作用子),这一术语表示起作用的单位,基本上符合通常指的基因或略小。一个作用子所包括的一段DNA与一个多肽链的合成相对应。平均的大小500-1500个核苷酸。
17.有互补作用,说明这两个突变分别属于两个基因位点,即非等位基因;如无互补作用,个体表现为突变型,则说明两个突变属于同一基因的不同座位,即等位基因。
18.有两个顺反子,分别为〔1,3,5,6〕和〔2,4〕。
19.本泽尔用大量的噬菌体rⅡ突变体对大肠杆菌B株成对进行双重感染。形成噬菌斑后,收集溶菌液(内含子代噬菌体),把它接到B株上,计算溶菌液中的总噬菌体数(因为两种rⅡ突变体rx、ry重组体r+r+和rxry都可以在B株生长),同时把溶菌液也接种到K12(λ)株上,计算野生型重组体r+r+数目,然后计算重组值:


用本法可以检出小到0.001%,即
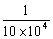 的重组值。利用大量rⅡ区内二点杂交的结果,可绘制出rⅡ区座位间微细的遗传图。本泽尔就是这样用经典的噬菌体突变和重组技术,对T4rⅡ区基因的微细结构进行了详细的分析。
的重组值。利用大量rⅡ区内二点杂交的结果,可绘制出rⅡ区座位间微细的遗传图。本泽尔就是这样用经典的噬菌体突变和重组技术,对T4rⅡ区基因的微细结构进行了详细的分析。
20.基因对遗传性状表达的作用,可分为直接的与间接的。如果它的最后产品是结构蛋白或功能蛋白,那么基因的作用可以直接影响到蛋白质的特性,从而表现出不同的遗传性状。例如人类的镰形红血球贫血症可以作为这方面的例证。研究证明,镰形红血球的血红蛋白是由一个正常血红蛋白基因(HbA)的两个不同的突变(HbS或HbC)引起的,即HbA→HbS或HbA→HbC。研究这三种血红蛋白HbA、HbS、HbC的氨基酸组成,发现这三者的不同,仅仅在于β链的第六位上有一个氨基酸的不同。在人的血红蛋白基因的密码中,仅仅改变其中的一个碱基,就可以直接引起它的最后产品——血红蛋白的性质发生改变,引起红血球镰形贫血症。
但是在更普遍的情况下,基因是通过酶的合成,间接地影响生物性状的表达。现以孟德尔的豌豆试验加以说明。高茎豌豆(TT)×矮茎豌豆(tt),其F1可以获得高茎豌豆(Tt)。已经研究证明,高茎的豌豆中含有一种促进茎部节间细胞伸长的物质——赤霉素,而矮茎豌豆中没有这种物质。赤霉素的产生需要酶的催化。高茎豌豆的T基因具有特定的核苷酸序列,它可以翻译成正常的促进赤霉素形成的酶,使它产生了赤霉素,细胞得以正常伸长,于是表现为高茎;矮茎豌豆的t基因则具有与T基因不同的核苷酸序列,它不能翻译成促进赤霉素形成的酶,因而不产生赤霉素,细胞不能正常伸长,而表现矮茎性状。这个例子清楚的说明,T与t基因控制株高的性状并不是直接的,而是通过指导赤霉素酶的合成而间接实现的。
21.1961年雅科和莫诺根据对大肠杆菌乳糖代谢的调控的研究,提出了一个操纵子模型。认为大肠杆菌的乳糖代谢所需要的三种酶的基因的转录受一个开关单位的控制,这种开关单位称操纵子。模型的内容图示如下:
当培养基内没有乳糖时,阻揭物接在操纵基因上,关闭了它所控制的操纵子,以阻止核糖核酸聚合酶的通过,使结构基因处于抑制状态,从而阻止了三种基因的转录和翻译。当培养基内加入乳糖后,细菌开始分解乳糖,分解的产物半乳糖便成为反阻遏的诱导物。诱导物与阻遏物相结合,便与操纵基因分离,从而打开了操纵子的开关,开放了核糖核酸聚合酶的通道,使结构基因活化,于是就开始了三种酶基因的转录和翻译,使三种酶量急剧增加。

22.从分子水平上看,一个基因位点还可以分成许多基本单位,称为座位。一个座位一般指的是一个核苷酸对,有时其中一个碱基发生改变,就可能产生一个突变。因此,突变就是基因内不同座位的改变。突变的方式主要有两种:一是分子结构的改变,如碱基替换和倒位;二是移码,如碱基的缺失和插入等。
23.紫外线作用于嘧啶,使得DNA同链上邻近的嘧啶核苷酸之外形成二聚体如胸腺嘧啶二聚体;或是将胞嘧啶脱氨成尿嘧啶,或是将水加到嘧啶的C4、C5位置上成为光产物。它可以削弱C—G之间的氢键,使DNA链发生局部分离或变性。实验证明,紫外线的作用集中在DNA的特定部位,显示了诱变作用的特异性。主要通过三种形式修复:
①光修复:由紫外线照射产生的
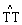 结构,被一种特殊的巡回酶,例如光激活酶所辨认,在有蓝色光波的条件下,二聚体被切开,DNA回复正常。这种经过解聚作用使突变回复正常的过程叫光修复。
结构,被一种特殊的巡回酶,例如光激活酶所辨认,在有蓝色光波的条件下,二聚体被切开,DNA回复正常。这种经过解聚作用使突变回复正常的过程叫光修复。
②暗修复,某些DNA的修复工作不需要光也能进行,例如,大肠杆菌中的UVrA突变体的修复过程由四种酶来完成:首先由核酸内切酶在
一边切开,然后由核酸外切酶在另一边切开,把TT和邻近的一些核苷酸切除;第三种酶(DNA聚合酶)把新合成的正常的核苷酸片段补上;最后由连接酶把切口缝合,使DNA的结构恢复正常。这类修复系统叫“暗修复”,或切除修复。
③重组修复:重组修复必须在DNA复制后进行,又称为复制后修复。这种修复并不切除胸腺嘧啶二聚体。修复的主要步骤如下:
1)含
结构的DNA仍可以进行复制,但子DNA链在损伤部位出现缺口。
2)完整的母链与有缺口的子链重组,缺口通过DNA聚合酶的作用,以对侧子链为模板由母链合成的DNA片段弥补。
3)最后在连接酶作用下以磷酸二酯键连接新旧链而完成重组修复。
24.遗传工程可分为狭义和广义的两种。狭义遗传工程即基因工程;广义的还包括细胞工程、染色体工程、细胞器工程等。习惯上所讲的遗传工程多指基因工程。基因工程是一种遗传操作,它采取类似于工程建设的方式,按照预先设计的蓝图,借助于实验室的技术,将某种生物的基因或基因组转移到另一种生物中去,使后者定向地获得新的遗传性状,成为新的类型。
从理论上看,基因工程是研究分子遗传学基本理论的一个重要方面,它既为细胞分化、生长发育、肿瘤发生等有关高等生物的基础研究提供了有效的实验手段,又为解决基因和基因组的精细结构、功能、控制机理问题提供了必要的分析手段。
从实践上看,基因工程的研究,将为解决农业、工业、医学等部门所面临的许多重大问题开辟了新的途径。如固氮基因的转移;提高工业发酵产品的产量和品质;治疗人的遗传疾病等。
25.一般来说,基因工程的施工程序大致分为四个步骤:第一为施工准备材料,即“目的”基因、载体和工具酶等;第二把目的基因与载体结合成重组DNA分子;第三把重组DNA分子引入受体细胞,建立分子无性繁殖系或称克隆;最后从细胞群体中选出所需要的无性繁殖系,并使外源基因在受体细胞中正确表达。
26.细菌细胞在大多数情况下能把渗入到细胞内的异源DNA分子毁灭,这称为限制。限制的能力来源于一种特殊的酶,这种酶称为限制性核酸内切酶。它是一种水解DNA的磷酸二酯酶,是遗传工程上的一种极其重要的酶,它可以把大的DNA分子切成所需要的基因或基因组的小片段,又可以把运载工具——质粒或病毒DNA切成适于携带基因组小片段的状态,以便形成重组DNA分子。因此,它的发现大大促进了遗传工程的发展。限制性内切酶可以作用于特定的碱基序列。例如在基因工程中常使用的限制性内切酶EcoRI,它所切断的部位是:
↓
 G A A T T C
G A A T T C
C T T A A G
↑
为了实现基因的转移,需要利用限制酶将“目的”基因安装在一个特定的运载工具——载体DNA上。在载体DNA的运载和保护下,被转移的基因可以高效地进入受体细胞,并在其中复制、扩增、转录和翻译,表达其一系列的功能,目前只有少数细菌质粒、λ噬菌体和某些动物病毒可以作为载体,但需要进一步改造,才能比较完善。在遗传工程中,最常用的运载工具是质粒。它们可以独立复制、稳定遗传,但是它的存亡与细胞的生存并无决定性影响。DNA的体外重组就是把“目的”基因牢牢地安装在载体上,使它们共价连接。DNA分子的重组,目前主要是限制性内切酶法。用同一种限制性核酸内切酶(如EcoRI)切割不同来源的DNA所产生的DNA片段,虽然区段和大小不同,但其单链末端(粘性末端)的顺序和长短都一样。因此,不同来源的DNA片段,可以通过(粘性末端)对应碱基间的氢键结合到一起,再经DNA连接酶的作用,将切口接合起来,成为一个完整的重组DNA分子。
27.植物体细胞杂交关键性的步骤有三个:
①原生质体的分离培养 首先选择适合的试验材料,一般用叶肉细胞和茎尖作材料较易成功,用培养的愈伤组织也能获得成功。其次是用果胶酶、纤维素酶、半纤维素酶以及崩溃酶等处理,分离出原生质体。然后将原生质体进行固体或液体培养。
②原生质体的融合 近年来最先进的融合技术是采用聚乙二醇(PEG)溶液处理混合的原生质体,
使之紧密粘聚,然后用高Ca++和高PH溶液处理。也可以采用电融合处理。
③杂种细胞的选择 异源原生质体经融合处理后,可以形成同源融合体、异源融合体和未经融合的原生质体,如何从这些大量的混合体中选择出杂种细胞,是体细胞杂交中一个极为重要的环节。目前已有三种较有成效的选择方法。第一个方法是营养选择法,第二个是互补选择法,第三个方法是异核体培养法。
28.最多有1500个突变子。实际检出的突变低于此数字。原因有四个:
①部分突变会造成简并密码子,因而表现不改变;
②不是所有的核苷酸都被转录;
③mRNA中不是所有核苷酸都被翻译;
④多肽前体在形成高级结构前有些氨基酸残基被切除。
29.两个。因为AUG是起始密码,同时也是甲硫氨酸密码,而UGA是终止密码。
30.可以,亚硝酸可以产生AT→GC,GC→AT的变化。
31.DNA: 3′ TAC CGT ACA GCA 5′
5′ ATG GCA TGT CGT 3′
mRNA: AUG GCA UGU CGU
反密码子: UAC CGU ACA GCA
肽链: 甲硫氨酸 丙氨酸 半胱氨酸 精氨酸
32.不可靠。因为双链DNA分子中A+G=T=C,所以A和G的总量应是50%。
33.假设最初一条染色体的DNA的两条单链都标记上,而新合成的链都没有标记上,则在生成的四条染色体上,有两条具有放射性。
34.双链DNA分子中的碱基组成应该是A=T、G=C。上列数据A≠ T、G≠ C,因此噬菌体ϕ174的DNA是单链的。
35.合成的(-)链中的41%T、17%G、25%C、17%A。因此合成的mRNA中含41%A、17%C、25%G、17%U。
36.a.属于mRNA;b.UAC; c.属于tRNA;d.ATG
37.大约是2100/3=700个。
38.2×108/(100×3)=666667。
39.43=64.
40.mRNA中G+C的含量是80%。
41.原来DNA序列为ATGGCATTAATTAGA,新DNA的序列为ATGCATTAAATTAGA;其中三联体1、4和5还是原来顺序,可以编码同样的氨基酸。
42.5-溴尿嘧啶是取代胸腺嘧啶的碱基类似物,常常同鸟嘌呤配对引起转换;
2-氨基嘌呤取代腺嘌呤和鸟嘌呤,能和胸腺嘧啶或胞嘧啶配对,DNA复制时造成碱基对的转换;
吖啶橙嵌入DNA分子中引起移码突变。
43.遗传密码具有以下特点:
①mRNA上密码子的排列是连续的。即连个密码子之间没有任何核苷酸将它们分开,因此阅读密码时,必须从一个特定的起点开始,一个密码子接一个密码子的阅读下去,直至终止信号为止,才不会发生阅读差错。
②通用性。所有生物不论是真核的还是原核的,均共用一套密码子,这说明地球上的生物有共同的起源。
③简并性。20种氨基酸由61个密码子编码,因此有的氨基酸有多个密码子编码。遗传密码的简并性对维持生物物种的稳定性具有重要的意义。
④变偶性。在密码子的三个碱基中,专一性主要取决于头两个碱基,第三个碱基比前两个专一性较小。因此与反密码子(在tRNA上)互补配对时,第三个碱基具有较大的灵活性,遗传密码的变偶性减少了密码阅读时的误差,增加了翻译的准确性。
44.在双链DNA中,A=T,G=C。如果A=15.1%,则T=15.1%,A+T=30.2%;G+C=100%-30.2%=69.8%,G=C=1/2×69.8%=34.9%。
45.一个密码子含三个核苷酸,这个细菌DNA分子90%的碱基所编码氨基酸数=

46.a. TTGATC b. GTTCGA c. ACGCGT d. ATGGTA。
47.a. T+C=0.46 b. T+C=0.54,A+G=0.46.
48.赖氨酸——甲硫氨酸——半胱氨酸——苏氨酸——缬氨酸
49.CAG-GAG-GTG-TCA-TAG。
50.a.5′ AUGCUAAC 3′ b.AUG c.AUG。
51.是以5′CTATTCGCACGTCACATTCGGGTA 3′转录而来的,其mRNA为
5′UACCCGAAUGUGACGUGCGAAUAG 3′。
52.

----A—A—C—A—G—A—C— G—G—T-----
----T—T—G—T—C—T—G—C—C—A-----
颠 颠 转颠
换 换 换换
53.碱基重复的结果将导致移码突变。
54.一些化合物,由于其结构与DNA分子中的某些碱基类似,因此被称之为DNA分子碱基类似物。它们常常能将掺入到DNA分子中去,好像DNA分子的正常组成成分,它们对DNA的复制影响不大,而在DNA复制时引起碱基配对上的差错,最终导致碱基对的替换,引起突变。例如5-溴尿嘧啶(5BU).当它以酮式状态存在时,分子结构与胸腺嘧啶基本相同,能替换DNA分子中的胸腺嘧啶。它掺入到DNA分子中后,通常情况以酮式状态存在,这是和胸腺嘧啶(T)一样,能与腺嘌呤(A)配对,但由于其上的溴原子的影响,他有时又以烯醇式状态存在于DNA分子中,这时它又能与鸟嘌呤(G)配对。当DNA复制到5-溴尿嘧啶时,其相对位置上将有鸟嘌呤与之配对。这样原来的A-T碱基对就会转变为G-C碱基对,导致突变的发生。其过程如图:

55.a.妨碍DNA某一成分的合成,引起DNA结构的变化。如6-硫基嘌呤能妨碍嘌呤的合成。
b.碱基类似物替换DNA分子中的不同碱基,引起碱基对的改变。如5-溴尿嘧啶可替换DNA分子中的T掺入DNA分子后又能与G配对,最后使A-T碱基对变为G-T碱基对。
c.直接改变DNA分子的某些特定结构。如甲基磺酸乙酯(EMS),可使DNA分子中的鸟嘌呤烷基化,形成7-烷基鸟嘌呤。7-烷基鸟嘌呤可以与T配对,从而产生GC→AT的转换。
d.引起DNA复制的错误。如2-氨基吖啶,能嵌入DNA双链中心的碱基之间,引起单一核苷酸的缺失或插入。
56.a.阻遏物和诱导物的区别:在分子组成上阻碍物是调节基因编码的一种蛋白质分子,诱导物是乳糖分解的产物半乳糖分子;在功能上,阻遏物与操纵基因结合,阻止转录进行;诱导物与阻碍物结合使之脱离操纵基因,转录得以进行。
b.操纵基因与调节基因的相似处在于它们都是由一段DNA分子组成,并且都参与对操纵子的调控。不同处在于操纵基因只能转录而不能翻译,调节基因即可转录又可翻译,产生调节蛋白;操纵基因通过与调节基因的产物的结合或释放来控制着整个操纵子的转录与否。
57.因为突变基因内部的另一个座位上的突变掩盖了原来座位的突变(但未恢复原来的密码顺序),故使突变的个体的表现型不变。
58.因为两条互补链转录的mRNA是不同的,所以带有不同的遗传信息。
59.因为一个顺反子内含有很多突变子。每个突变子产生的效应可以完全不同。例如有的突变仅造成简并密码子,并不影响生物体内蛋白质的结构和功能。这样生物体的表现型就不会发生改变。也有的突变体内蛋白质不能生成,或者失去正常功能,从而影响有机体的发育和生长,出现致死效应。
60.顺反测验就是根据顺式表现型和反式表现型来确定两个突变体是否属于一个基因或顺反子。实际上,顺式排列只是作为对照,一般并不进行测验,因为它的表现型是野生型。所以测验实质上是反式测验。如反式测验是野生型,说明这两个突变分别属于两个基因位点,既非等位基因;如表现为突变型,则说明两个突变属于同一基因的不同座位,即等位基因。
61.EcoRI切断的部位是:
↓
G A A T T C
C T T A A G
↑
HindIII切断的部位是:
↓
A A G C T T
T T C G A A
↑
这两种限制性内切酶所切断的DNA分子多数可以产生粘性末端,即在酶解时,DNA双链不在同一地方断开,因而产生的片段两端都带有数个碱基的单链末端,这两个单链末端带有互补的碱基配对顺序,可以互相自动接合成为环状DNA。
62.不同,因为A=T≠G=C
63.反转录酶可以以RNA为模版,合成DNA。迄今不仅在几十种由RNA致癌病毒引起的癌细胞中发现反转录酶,甚至在正常细胞,如胚胎细胞中也有发现,这一发现增加了中心法则中遗传信息的原有流向,丰富了中心法则的内容。反转录酶的发现不仅具有重要的理论意义,而且对于遗传工程中基因的酶促合成,以及致癌机理的研究都有重要的作用。
64.只有mRNA有显著差异。
65.a.双链DNA; b.RNA; c.单链DNA
66.第二个样品比第一个样品含有较高的G-C碱基对。因为G-C碱基对由三个氢建相连,而A-T碱基对由二个氢建相连。所以G-C碱基对含量高的DNA分子变性温度也高。
67.大肠杆菌的复制速度为90000×3.4
/min=30.6 μm/min,所以染色体长度为1100μ m的大肠杆菌复制一次需用1100 μm/30.6=36min。(1μ=10000
)
68.由于起转化作用的是甲细菌的DNA,所以用DNA酶处理失活甲细菌后,可使甲细菌的DNA由于DNA酶的作用而被降解,因此用DNA酶处理的失活甲细菌再与活性乙细菌混合一块处理小鼠,小鼠将不会染肺炎而死亡。
69.G+C/A+T
70.1/0.5=2;0.5
71.A/G=2;A/C=2;A/T=1
72.属两个基因:a、b属于一个基因,c为另一个基因。
二、判断对错
1.×;如果含A-T碱基对多则解链温度低,如果含G-C碱基对多则解链温度高。
2.√
3.×;包括遗传密码的转录和翻译这两个步骤。
4.×;是RNA
5.×;反密码子是3′AUG5′。
6.√
7.×;说明属于两个基因
8.√
9.×;细菌转化现象是1928年由格里菲斯(Griffth,F.)发现的,而重复出这种现象并证明转化因子的是阿委瑞(Avery,O.T.)
10.×;亚硝酸在一定条件下,可将鸟嘌呤和胞嘧啶氧化成为次黄嘌呤和尿嘧啶。
11.×;两种酶都可将DNA分子切成粘性末端。
三、选择答案
1.b
2.a
3.b
4.c
5.b
6.a、c、d
四、填空
1.不同座位
2.调节基因
3.松弛型;严紧型
4.瓦特森(Watson,J.D.);克里克(Crick,F.H.C.)
5.60;40;80
6.野生型;这两个突变分别属于两个基因位点,既非等位基因;两个突变属于同一基因位点,即等位基因。
7.20
;3.4
;34
;10
8.大肠杆菌
9.核酸内切酶;核酸外切酶;DNA聚合酶;DNA连接酶
10.营养选择法;互补选择法;异核体分别培养法
11.限制
12.转化;转染
13.结构基因;操纵基因;调节基因
14.座位
15.“目的”基因;载体;限制性核酸内切酶
五、名词解释
基因文库:即把一种生物的全部基因像图书馆藏书一样储存在受体细胞里。
启动子:DNA分子上能与RNA多聚酶结合而使转录开始的区段。
终止子:染色体上转录终止的部位。
限制:生物细胞具有防御异源遗传信息进入的手段,异源DNA分子渗入细胞后,在许多情况下遭到毁灭,这个过程称为限制。
转染:把重组噬菌体或重组病毒DNA引入受体细胞,称传染。
半保留复制:DNA自身的复制方式,复制时DNA双链解开并以每一单链为模板来形成另一对应的新链。
遗传密码:生物蛋白质合成的密码。是遗传信息的单位。由构成核酸的四种不同核苷酸的不同组合所代表。每一密码子由核酸分子中三个相连核苷酸所组成,决定一个氨基酸,所以称三联体密码。
简并:一个氨基酸由一个以上的三联体密码所决定的现象。
翻译:指mRNA携带者转录的遗传密码附着在核糖体上,把由转移核糖核酸(tRNA)运来的各种氨基酸,按照mRNA的密码顺序,相互连接起来成为多肽链,并进一步折叠起来成为立体蛋白质分子。
转录:遗传信息从DNA到mRNA的转移,亦即以DNA为模版由四种核糖核苷三磷酸合成mRNA的作用。
多核糖体:在蛋白质合成过程中,一个mRNA要同两个以上的核糖体结合起来,形成一串核糖体,称为多核糖体。
突变子:它是性状突变时,产生突变的最小单位。一个突变子可以小到只是一个核苷酸对。
重组子:发生性状的重组时,可交换的最小单位称为重组子。据微生物重组的精细研究,一个交换子只包含一对核苷酸。
顺反子:这一术语表示一个起作用的单位,基本上符合通常指的基因或略小。
重组基因:指同一段DNA的编码顺序,由于阅读框架的不同或终止早晚的不同,同时编码两个以上基因的现象。
隔裂基因:指一个基因内部为一个或更多的不翻译的编码顺序,及内含子所隔裂现象。
基因内抑制:指突变基因另一座位上的突变掩盖了原来座位的突变(但未恢复原来的密码顺序),使突变型表现野生型性状。
无义突变:一对碱基的改变而使某一氨基酸的密码子变为一个终止密码子的的突变。
误义突变:导致某种蛋白质分子中某一氨基酸为另一氨基酸所替代的突变。
移码突变:一对或少数几对邻接的核苷酸的增加或减少造成这一位置后一系列编码发生移位错误的突变。
碱基替换:一个碱基对被另一碱基对替代。
转换:碱基替换时,嘌呤由嘌呤代替,嘧啶由嘧啶代替叫转换。
颠换:碱基替换时,嘌呤由嘧啶代替,嘧啶由嘌呤替换叫做颠换。
跳跃基因:指在染色体上可以转移的基因。
调节基因:控制结构基因转录和翻译的基因。
结构基因:指决定蛋白质(包括酶)结构的基因。
操纵基因:控制结构基因转录的开关位点。
位点:从细胞水平上理解,基因相当于染色体上的一点,称为位点。
座位:从分子水平上看,一个基因位点开可以分成许多基本单位,称为座位。一个座位,一般指的是一个核苷酸对。
基因间抑制:指控制翻译机制的抑制基因,通常是tRNA基因发生突变,从而使原来的无义突变,误义突变或移码突变恢复成野生型。
限制性内切酶:一类能作用于外源DNA分子双链特定碱基顺序的特定位点并切割产生具有粘性末端的DNA片段的酶。各种限制性内切酶有专一性识别位点。
克隆:通过有丝分裂,从一共同的细胞或生物繁殖得到一群基因型完全相同的细胞或生物。
粘性末端:DNA双链分子末端的一小段单链部分。由于和同一分子的另一端或另一分子的一端的单链部分具有互补碱基,所以能通过互补碱基之间的氢建的形成并通过连接酶的作用连接起来,使同一分子的两端连接成为双链从而出现环状分子。或使两个DNA分子连接成为一个。粘性末端可通过限制性酶的处理而得到,噬菌体λ等具有天然的粘性末端,使它感染寄主后能形成环状DNA。
Hela细胞:指1953年从一名美国女孩Helen Lane的子宫颈上取出培养的癌细胞系。
异核体:含有属于不同基因型的核的细胞,也指由这些细胞所构成的个体。
互补作用:两个带有不同突变基因的染色体处在同一细胞中,由于相对的野生型的互相补偿而使表现正常化的作用。
操纵子:由操纵基因以及紧接着的若干结构基因所组成的功能单位。其中的结构基因的转录为操纵基因所控制。
第九章细胞质遗传
一、回答问题
1.相同处:两者都是按半保留方式进行自我复制,一般情况下都是通过转录mRNA,翻译蛋白质的方式控制性状的表达。
不同处:
①存在位置不同,核基因载于核内染色体上,细胞质基因载于细胞质的细胞器或其他细胞质颗粒上。
②遗传方式不同,核基因表现孟德尔式遗传,细胞质基因表现母性遗传。
③在作用上,决定个体发育的方向和模式方面,核基因占主导地位。
2.a.绿色;b.白色;c.绿色、白色、花斑。
3.果蝇对CO2反应能力的遗传是受细胞质基因决定的。
4.相同点:正反交的遗传表现不同,都表现母本性状。
不同点:母性影响是由于母本细胞核基因的产物积累在卵细胞中,从而使杂种F1代的性状表现与雌亲相似的遗传现象。故这种遗传现象决定于受精前母体卵细胞状态,不决定于细胞质基因。而细胞质遗传则是由母本细胞质基因作用的结果,其特点是非孟德尔式遗传。
5.如为性连锁遗传,其正反交结果不同将与性别相联系。
如为细胞质遗传,则某性状只有在母本表现时才能遗传给后代,连续近亲繁殖时,性状不分离也不消失。
如为母性影响遗传,则经连续近亲繁殖后,F3代会出现性状的分离。
6.这种草履虫的遗传组成为:kk+卡巴粒。由于细胞核中不含K基因,细胞质中的卡巴粒不能持续存在,经几次细胞分裂后逐渐减少,最后消失,成为敏感型的个体。
7. P: ♀dd左旋×♂DD右旋
↓
F1: 左旋Dd
↓
F2:DD右旋 Dd右旋 Dd右旋 dd右旋
8.将具有目的性状的个体作为母体。
9.a.品系A基因型为KK,有卡巴粒;品系B为KK,无卡巴粒;品系C为kk,无卡巴粒。
b.品系D表现型为放毒型,基因型为Kk,细胞质中有卡巴粒。
10.两个基因型为Kk草履虫可以以三种方式进行单倍体的细胞核融合,即:K×K、K×k、k×k,比例分别为:
 、
、
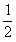 、
、
。K×K产生的两个个体是KK和KK,K×k结合产生的后代是Kk和Kk,k×k产生的后代是kk和kk。因此各种后代的概率为:a.KK和KK为
;b.KK和Kk为0;c.KK和kk为0;d.Kk和Kk为
;e.kk和kk为
。
11.a. Kk(卡巴粒)× kk
↓
Kk(卡巴粒) kk(卡巴粒)Kk kk
↓↘ ↓ ↓↘ ↘
↓ ↘ ↓ ↓ ↘ ↘
↓ ↘ ↓ ↓ ↘ ↘
KK(卡巴粒)
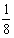 kk
kk
kk
KK
kk
kk
经过多次分裂后放毒型(KK+卡巴粒)占1/4。
b. Kk(卡巴粒)× kk
↓
Kk(卡巴粒) kk(卡巴粒)
↓↘ ↓
↓ ↘ ↓
↓ ↘ ↓
KK(卡巴粒)
kk
kk
12.a.全部为敏感性;b.全部为抗性。
13.决定质体性状的遗传基础与细胞质基因有直接联系。O1细胞质中必有绿色质体基因,O2细胞中必有黄色质体基因。
14.除了DD可以确定右旋外,其它基因型的表型型无法确定。因为它们的表型是受母本基因所决定的。
15.a.全部正常;b.全部正常。
16.不育系S(rr),保持系N(rr),恢复系S(RR)或N(RR),恢复性杂合体S(Rr)或N(Rr)。
17.遗传组成:前者是核内染色体上不育基因决定的。后者是由染色体易位造成的。表现形式:前者花粉完全败育,但胚囊可育。后者无论花粉还是胚囊均为半不育。
18.孢子体不育是指花粉的育性受孢子体(植株)基因型所控制。而与花粉本身所含基因无关。如果孢子体的基因型为rr,则全部花粉败育;基因型为RR,全部花粉可育;基因型为Rr,产生的花粉有两种,一种还有R,一种还有r,但两种花粉都可育。配子体不育是指花粉的育性直接受雄配子体(花粉)本身的基因所决定的。如果配子体内的核基因为R则该配子可育;如果配子体内的核基因为r,则该配子不育。
19.a.不育系S(rr)×恢复系N(RR)→S(Rr)
b.不育系S(rr)×保持系N(rr)→S(rr)
c.不育系S(rr)×恢复性杂合体S(Rr)→S(Rr)、S(rr)
20. A: S(rr)×B: N(rr) C: S(rr)×D: N(RR)
↓ ↓
AB: S(rr) ×CD:S(Rr)
↓
S(Rr),正常可育:
S(rr),雄性不育
21.杂合的配子体质核不育型S(Rr)
22.核不育的不育性状是由细胞核内一对隐性基因控制,纯合体(msms)表现为雄性不育,这种不育性为显性基因MS所恢复,杂合体(Msms)后代呈简单的孟德尔式分离。通常情况下不能使不育性保持。因此,其在生产上的利用受到了很大限制。
23.a.根据雄性不育性的败育发生过程,可分为孢子体不育和配子体不育两种类型;
b.胞质不育基因的多样性与核基因的对应性,即同一植物内可以有多种质核不育类型;
c.不育性既有单基因控制的,也有多基因控制的。
24.亲本不育系为配子体不育。
P:S(rr)×N(RR) S(rr)♀ S(Rr)♂
不育系↓ 恢复系 ↓ ↓ ↘
F1: S(Rr) r R r(不育)
↙ ↘ ↘ ↙
R r(不育) S(Rr)一半花粉可育
↓
F2:S(RR)可育,S(Rr)一半花粉可育
25.a.3[S(R1_r2r2)]半不育:1[S(r1r1r2r2)]不育
b.3[S(R1_R2_)]可育:1[S(R1_r2r2)]半不育
c.S(r1r1R2R2)全部为半不育
d.9[S(R1_R2_)]可育:3[S(R1_r2r2)]半不育:3[S(r1r1R2_)]半不育:1[S(r1r1r2r2)]不育
26.在雄配子体中两者表现相同,都为半不育;在雌配子体中前者为完全可育,后者仍未半不育。
27.a.由于自交不亲和复等位基因间的相同基因的拮抗作用;
b.同源三倍体染色体分离的不均衡性; c.易位杂合体;
d.质核互作的孢子体不育基因; e.无配对的同源染色体而导致染色体分离的紊乱。
28.a.隐性核雄性不育基因引起,特点:服从孟德尔遗传规律。
b.核不育基因与胞质不育基因共同决定。特点:可用恢复系恢复其育性,也可用保持系保持其不育性。
c.倒位杂合体引起花粉败育。特点:部分不育。
d.缺失杂合体引起的花粉败育。特点:部分不育
e.易位杂合体引起的花粉败育。特点:半不育
f.同源三倍体。特点:多数植株表现为高度不育
g.单倍体植株。特点:染色体不能配对,加倍后变为可育
29.同一植物内可以有多重质核不育类型,但是由于胞质不育基因和核基因的来源和性质不同,在表现特征和恢复性反应上往往表现明显的差异,即胞质不育基因的多样性与核育性基因的对应性。对于每种不育类型,都需要某一特定的恢复基因来恢复。
30.该不育系属孢子体质核互作雄性不育型。不育性与两对具有互补关系的核基因有关。当两个育性基因R1和R2同时存在时表现为花粉可育,否则为花粉败育。
不育系S(r1r1r2r2)×N(R1R1R2R2)恢复系
↓
♂S(R1r1R2r2)×♀S(r1r1r2r2)
↓
S(R1r1R2r2) S(R1r1r2r2) S(r1r1R2r2) S(r1r1r2r2)
可育 不育 不育 不育
31.该不育性状是受显性单基因控制的核不育型。
32.A是这个不育系的恢复系,B是它的保持系。C对B是显性核不育,C对A是隐性核不育。不育基因Msc与恢复基因Msa和保持基因Msb是位于同一基因位点上的三个复等位基因。其显隐性关系为Msa>Msc>Msb。杂交过程如下:
(C不育系)Msc Msc×Msb Msb(B保持系)
↓
F1 MscMsb(不育)


BC1:MscMsb,Msb Msb F2:Msc Msc,MscMsb,Msb Msb
1不育株:1不育株 1不育 :2不育 :1可育
(C不育系)Msc Msc×Msa Msa(A恢复系)
↓
F1 MscMsa(可育)


BC1:MsaMsc,Msa Msa F2:Msa Msa,MsaMsc,Msc Msc
全部可育 1可育 :2可育 :1不育
33.配子体不育。
34.首先以已有的不育系为母本,某个优良的品种为父本杂交,然后再用这个优良品种为轮回亲本进行多代回交。回交过程中注意选择既具有雄性不育的特征,又具有优良品种的性状。多代回交后,可获得理想的不育系。这个优良的亲本品种就是它的保持系。通常采用测交筛选法选育理想的恢系。方法是,以现有的优良品种、品系为父本,与不育系测交,在杂种一代中选择具有恢复性能强的优良组合,其父本就是这个不育系的恢复系。也常用回交转育法选育恢复系。方法与理想不育系的选择基本相同。利用不育系与现有恢复系杂交,再用优良的品种、品系为轮回亲本进行多代回交。在回交后代中选择出恢复性能强、性状优良的个体即为优良的恢复系。
35.①质核互补控制假说:这个假说认为只要质、核双方有一方携带可育的遗传信息,无论是N或R,都能导致花粉正常发育。R可以补偿S的不足,N可以补偿r的不足。只有S与r共存时,由于不能相互补偿,所以表现为不育。
②能量供求假说:认为植物的育性与线粒体的能量转化率有关。这个假说是假定供能水平的高低取决于mtDNA,而耗能水平的高低取决于核基因。进化程度低的野生种或栽培品种的线粒体能量转化率低,供能低,耗能也低,供求平衡,所以雄性可育。反之,进化程度高的栽培品种,线粒体能量转化率高,供能高,耗能也高,供求平衡,因此雄花育性正常。如果低供能的作母本,高耗能的作父本,得到的质核杂种由于能量供求不平衡,因而表现雄性不育。如果高供能的做母本,低耗能的做父本,由于杂种的供能高而耗能低,因而育性正常。
③亲缘关系假说:认为双亲间的亲缘关系差距越大,杂交后的生理不协调程度也越大。这种不协调达到一定程度,就会导致花粉的败育。
二、判断对错
1.×;只有一种左旋类型。
2.×;玉米的条斑是由隐性的核基因(ijij)引起的胞质基因变异,变质的胞质基因引起玉米的条斑性状。
3.×;K基因的作用是使卡巴粒在细胞质中持续存在。
4.√
5.√
6.×;核不育性不易受环境条件的影响,而质核不育性则易受环境条件的影响。
7.×;mtDNA是线粒体DNA,ctDNA是叶绿体DNA。
8.×;杂种后代呈现株间育性分离的雄性不育是孢子体不育,而杂种植株呈现株内花粉育性分离的则是配子体不育。
9.√
10.×;前者只是花粉表现为半不育,而后者胚囊和花粉都表现半不育。
11.√
三、选择正确答案
1.b;‚c;ƒc;④d。
2.a;‚b。
3.Ⅰ、b;Ⅱ、a;Ⅲ、c;Ⅳ、b;Ⅴ、d。
四、填空
1.孢子体;配子体
2.不育性能好;一致;恢复性能强;配合力高;农艺性状好
3.细胞质不育基因和细胞核不育基因共同;孢母细胞染色体分离的不均衡性,即单倍体
4.核基因;花粉母细胞减数分裂;彻底;明显可分;保持系。
5.多;营养;二;遗传
6.核;质核互作
7.恢复系;N(RR)或S(RR);保持系;N(rr);质核互作
8.胞质不育基因;核不育基因
9.细胞核基因和全部或绝大部分细胞质基因;细胞核基因
10.质核不育基因互作;线粒体基因组;叶绿体基因组;卡巴粒

五、名词解释
共生体:由外部进入细胞的共生成分(微生物),它能与寄主细胞共同存在,并使寄主细胞具有某些特有的性状,但并非是细胞生命活动所必须的物质。
细胞质基因组:所有细胞器和细胞质颗粒中的遗传物质。
核遗传:由染色体基因组所控制的遗传现象和遗传规律。
细胞质遗传:由细胞质基因所决定的遗传现象和遗传规律。
母体遗传:同细胞质遗传。
母性影响:由于卵细胞质中存在母体核基因的某些代谢产物,使子代的性状并不受本身的基因型所决定,而表现与母体相似性状的遗传方式。
前定作用:由受精前母体卵细胞装袋决定子代性状表现的现象。同母性影响。
延迟遗传:同前定作用,母性影响。
自体受精:同一个体产生的雌雄配子的结合过程。
雄性不育遗性:植株的雄蕊发育不正常,不能产生正常功能的花粉,但是它的雌蕊发育正常,能接受外来正常花粉而受精结实。
雄性不育系:雄蕊发育不正常,失去受精能力,雌蕊发育正常,能接受外来花粉进行受精的植物品系。简称A系。
保持系:指给不育系授粉后能使其后代仍保持雄性不育性状的父本品系。简称B系。
恢复系:指给不育系授粉后能使其后代雄蕊发育恢复正常的父本品系。简称C系。
核不育型:核内染色体上基因所决定的雄性不育类型。
质-核不育型:由细胞质基因和细胞核基因互作控制的雄性不育类型。
孢子体不育:指花粉的育性受孢子体(植株)基因型所控制,而与花粉本身所含基因无关。
配子体不育:指花粉的育性直接受雄配子体(花粉)本身的基因型所控制。
同型保持系:除了具有雄性可育性状外,其它性状完全与其不育系相同的一种保持系。
第十章 遗传与发育
一、回答问题
1.个体发育有两个特点:一是个体发育的方向和模式由受精卵中的基因型决定,由基因型和环境条件共同作用形成个体的各种性状;二是受精卵分裂到一定阶段,细胞开始分化,产出具有不同形态结构和生理功能的组织和器官,使其细胞核内仍具有相同的遗传物质。
2.在个体发育过程中,细胞核和细胞质是相互依存,不可分割的,但起主导作用的应该是细胞核。没有细胞核只有细胞质的细胞在短时间内,还可以进行一些生长和分裂,但最终必死亡。这是因为蛋白质的合成需要细胞核不断供应mRNA,没有细胞核就没有mRNA的来源,没有mRNA,蛋白质合成就会停止。若没有细胞质只有细胞核,这种核可以在体外培养,但到目前为止,只能做短期培养。这是因为核内DNA的复制,mRNA的转录以及tRNA、rRNA的合成都需要原料和能量,这些原料和能量总得从细胞质来。所以只有细胞质和细胞核一起协同作用,才可使生命通过生长和繁殖而维持下去,可见,在这方面两者是相辅相成的,缺一不可。
3.海胆卵分裂为四个细胞时,每个卵裂细胞都能发育成小幼虫,因为海胆受精卵的第一、二次分裂都是顺着对称轴的方向进行的,各个卵裂细胞中不仅细胞核的内容一致,而且细胞质的成分也是完全相同的。但分裂为八个卵裂细胞后再分开,就不能发育成小幼虫,因为第三次卵裂方向与对称轴垂直,虽然细胞核仍相同,但由于细胞质的不等分裂,这八个细胞的细胞质及其中的细胞器却不相同,所以不能发育成幼虫,说明细胞质对细胞分化起着重要作用。
4.在个体发育过程中,从受精卵到胚胎,从营养成长到生殖生长,随着发育时期的推进,相应的性状在各个阶段有节奏的发生,各部分细胞分化成不同的形态特征和生理特征,这就是个体发育的阶段性。个体发育过程包括一系列相继的发育阶段,这些阶段按预定的顺序依次接连发生,上一阶段趋向完成就启动下一阶段的开始,相互不能超越,这就是个体发育的连续性。是生物体发育最显著的特征之一,产生的原因在于个体各部分细胞的基因不是同时表达,而每个细胞的全部基因也不是在个体发育过程中都始终一贯地表达。
5.个体发育阶段性转变的过程,实质上是不同基因的被激活或被阻遏的过程。在发育的某个阶段,某些基因被激活而得到表达,另一些基因则处于被阻遏或保持静止的状态。在发育的另一阶段,原来被阻遏的基因因激活而表达了,原来表达的基因却被阻遏。
6.组蛋白与DNA分子相结合能阻止转录的进行,而非组蛋白能激活受阻遏的DNA分子的转录。
7.分化——广义的说就是一种类型的细胞在形态上、生理上以至生物化学上转变成另一种类型的细胞,这种变化是不可逆的。高等植物的分化过程从形态上、生理上可分为三个阶段:从受精卵分化为胚,称为胚胎发生。‚胚进一步生长和分化形成营养体称为营养生长。ƒ营养体分化成花,称之为生殖生长。
去分化(脱分化)——离体的植株器官、组织或细胞在人工培养基上经过多次细胞分裂而失去原来的分化状态,形成无结构的愈伤组织或细胞团。
再分化——离体培养的植物细胞和组织,可以由分化状态再度分化形成另一种或几种类型的细胞、组织或器官甚至最终长成完整植株。
由于合子通过一系列的有丝分裂所产生的各个细胞,都具有全套的遗传信息以及传递信息的转录和翻译的功能,因而都具有与合子相似的发育新个体的功能。但在个体发育过程中,由于各个细胞受发育阶段所在组织器官环境的约束,只能表现一定的功能,而不能表达全部功能。一旦脱离了这种约束,成为游离状态,并且得到必须的营养和激素,就可能去分化而恢复其遗传全能性,具有类似合子的功能。
二、填空
1.5;Hbα,Hbβ,Hbγ,Hbδ,Hbε
2.营养核;稠密;生殖核
3.组蛋白;阻遏
三、名词解释
个体发育:高等生物从受精卵开始,经过一系列细胞分裂和分化,长成新的个体。这个过程通常称为个体发育。
细胞全能性:指个体某个器官或组织已经分化的细胞再生成完整个体的遗传能力。
分化:使一个生物的形态和功能不断地趋向于不同的一系列变化。
去分化:离体的植株器官、组织或细胞在人工培养基上经过多次细胞分裂而失去原来的分化状态,形成无结构的愈伤组织或细胞团。
第十一章 遗传和进化
一、回答问题
1.生物进化的理论可归纳为两个,一是拉马克的获得性遗传说,另一个是达尔文的自然选择学说。
前者认为,生物生存的外界条件的改变是引起遗传特征变异的根本原因。因外界坏境条件对生物的影响有两种形式;对于植物和低等动物,这种影响是直接的。对于具有发达神经系统的高等动物则是间接的。当外界环境条件改变时,首先引起动物习性和行为的改变,然后促使某些器官使用的加强活减弱。这样,由于用进废退和获得性的遗传,生物逐渐到得发展。后者虽然同意拉马克的观点,但把选择的作用提到了首位。认为自然选择和人工选择是生物进化的主要动力。达尔文认为,不论在自然界或在栽培条件下,变异是广泛存在的,这些变异许多是可以遗传的。通过多代对这些变异的自然选择人工选择,从而使之向更适应于环境条件的方向发展。
2.随着地球上自然条件的变化,生物经历着一个由简单到复杂,由低级到高级的进程,这就是生物的进化。按着进化论的观点,生物进化是通过对生物体可遗传变异的选择而实现的。而产生可遗传变异的原因是突变和基因重组,而突变和基因重组正是遗传学所研究的范畴。因此可以说遗传学是研究生物进化的理论基础。
3.群体大;个体间能自由交配;无外界因素干扰。
4.有害的隐性伴性基因比有害的常染色体隐性基因表现出来的频率高,因此容易受到自然选择的作用。
5.由于在有限群体内取样的误差而产生的基因频率的随机波动称为遗传漂移。这种作用可将群体中一些中性或无任何适应价值的性状保留下来。
6.在一个完全随机交配的群体内,如果没有其它因素干扰,则基因频率及基因型频率常保持一定,各代不变,这就是群体遗传平衡定律。其要点是:第一,这个群体必须是随机交配的;第二,无其它因素干扰;第三,达到平衡状态时,这个群体的基因频率和基因频率的关系是;D=p2, H=2 pq, R=q2。影响群体遗传平衡的因素有;突变、选择、迁移和遗传漂移。
7.根据哈德—魏伯格定律,在一个随机交配的平衡群体中,各代的基因频率和基因型频率保持不变,因此,品种的典型性能代代相传不变。
8.不,根据哈德-魏伯格定律,如果没有其他因素的干扰,不论是显性基因还是隐性基因,群体中的基因频率逐代不变。
9.随机交配。
10.突变是物种形成的主要因素,它为新物种的形成提供原始材料。隔离是物种形成的必要条件,那些发生了变异的个体或群如果不同原群体隔离开,仍然任其自由交配,那么形成的新基因和原群体中的基因进行交流,新的性状在群体中很快消失,而不能形成新物种。
11.凡能杂交而产可育后代的群种为一个物种。物种形成有两种不同的方式:
渐变式:在一个长时期内,旧的物种逐渐累积变异而演变成为新物种。其中又分为继承式和分化式两种。继承式是指一个物种可以通过逐渐累积变异的方式经过悠久的地质年代,由一系列中间类型过渡到新种。例如马的进化历史。分化式是指一个物种的两个或两个以上的群体,由于地理隔离或生态隔离而逐渐分化成两个或两个以上新种。例如棉属中的一些种的变化。
‚爆发式:不一定需要悠久的历史,在较短的时间内即可形成新种。一般也不经过亚种,而是通过染色体变异或基因突变以及远缘杂种的染色体加倍,在自然选择的作用下逐渐形成新种。例如普通小麦的形成。
12.多倍体是植物界新种形成的重要途径。通过不同物种的染色体合并和加倍,成为多倍体而形成新种。新种可以在较短的时间内形成,一般也不经过亚种阶段。可以在自然条件下自发进行,也可人工创造。所谓爆发式物种形成的主要途径之一就是通过多倍体形成来完成的。
13.将两种植物杂交,如果能够产生可育的杂种后代,为同一物种。否则为不同物种。
14.根据公式

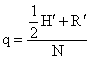
所以
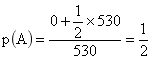
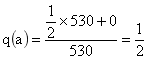
15.群体‚处于平衡状态。
因为
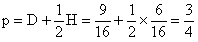
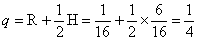
平衡状态时应有D=p2 H=2pq R=q2
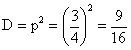
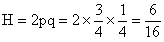
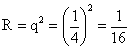
16.
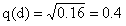
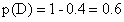
杂合体的比例:
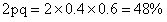
17.设亲代中l的基因的频率为
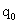 ,第二代中1的频率为
,第二代中1的频率为
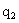 ,L的频率为
,L的频率为
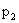 。
。
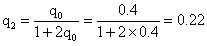
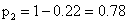
第二代中表型正常个体中携带1基因的个体的比例为:
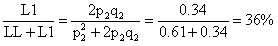
18.
 ,
,
 ,
,
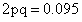
19.
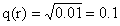
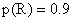
红花基因频率
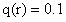
白花群体中纯合体的比例为:
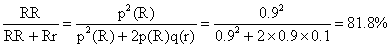
白花群体中杂合体的比例为:
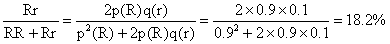
20.群体中男性色盲患者的频率就是色盲基因的频率即:
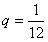
a.女性中色盲的比例为

b.群体中色盲女人的比例为

21.
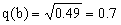
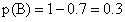
22.红色斑点基因频率为0.1,黑色斑点基因频率为
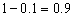
黑色牛中纯合体的比例
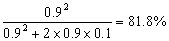
黑色牛中杂合体的比例

23.依题意可知
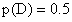 ,
,
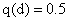 。连续自交5代时,各种基因型频率及个体数是:
。连续自交5代时,各种基因型频率及个体数是:
 ,共有
,共有
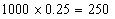 只
只
 ,共有
,共有
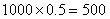 只
只
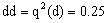 ,共有
,共有
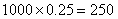 只
只
从第6代到第8代连续两次选择,此时基因d的频率变为:

D基因的频率为
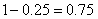
24.O型血的比例为:

A型血的比例为:

B型血的比例为:
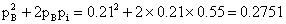
AB血型的比例为:

25.

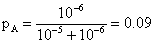
26.
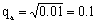
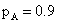 ,
,
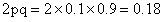 。携带这种基因的正常株的比例为18%。
。携带这种基因的正常株的比例为18%。
27.
 ,
,
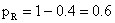 ,
,
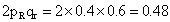 。两种配子相互结合的机会为48%。
。两种配子相互结合的机会为48%。
28.a.
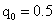 ,
,
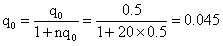
b.

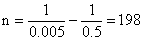 (代)
(代)
29.
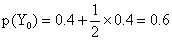
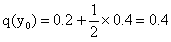
a.
 代基因型频率:
代基因型频率:
 ,
,

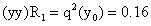
根据哈德—魏伯格定律要点:在一个大群体中,不论基因频率和基因型频率如何,只要一代的随机交配,这个群体就可以达到平衡。所以F1代基因频率为
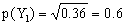 ,
,
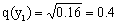 。
。
 代的基因型频率和基因频率与F1相同。
代的基因型频率和基因频率与F1相同。
b.
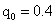 ;
;
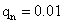 ;
;
 (代)
(代)
30.设混合前群体Ⅰ的隐性基因频率为q1、群体Ⅱ的隐性基因频率为q2。混合前隐性基因纯合体频率的平均值为
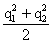 ,混合后隐性基因纯合体的频率为
,混合后隐性基因纯合体的频率为
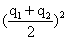 。
。
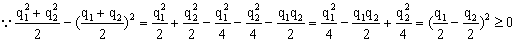
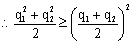
31.设白化基因的频率为q,
 ;群体中正常植株产生携带白化苗基因的配子的比例为:
;群体中正常植株产生携带白化苗基因的配子的比例为:
 ,因此正常植株的后代出现白化苗的频率为:
,因此正常植株的后代出现白化苗的频率为:
 。
。
32.a.合并后基因型dd的频率为
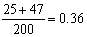 。基因d的频率为0.6。
。基因d的频率为0.6。
b.根据哈德—魏伯格定律,自由交配的群体中各代基因频率不变,所以下一代中基因d的频率仍为0.6。
c.
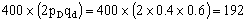
33.基因型e/e是完全致死的,等位基因e的频率在选择作用下依下述公式降低:
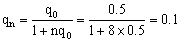
34.a.结合后群体中引起这种疾病的基因频率:
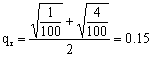
b.结合后群体下一代此病患者的比例为:
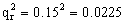
c.结合前患病平均比例为
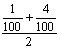 ,结合后患病的比例为0.0225。结合后比结合前的平均患病率低
,结合后患病的比例为0.0225。结合后比结合前的平均患病率低
 。
。
35.在红芽鞘隔离区内,绿色基因频率
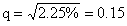 ;红色基因频率
;红色基因频率
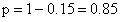 。因此,携带绿色基因的个体比例为
。因此,携带绿色基因的个体比例为
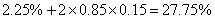 。在绿芽鞘隔离区内携带红色基因的个体比例共占13%。所以红芽鞘隔离区的混杂度高。
。在绿芽鞘隔离区内携带红色基因的个体比例共占13%。所以红芽鞘隔离区的混杂度高。
36.迁入率
 ;迁入的群体中A基因的频率
;迁入的群体中A基因的频率
 ;原群体中A的频率
;原群体中A的频率
 。经过一个世代的交配后,A的频率为:
。经过一个世代的交配后,A的频率为:
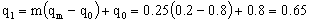
基因A的变化
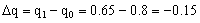 ,即比迁入前基因A的频率下降了0.15。
,即比迁入前基因A的频率下降了0.15。
二、判断对错
1.×;还必须无外界因素干扰。
2.×;
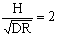
3.×;混杂度为:

4.√
5.√
6.√
三、填空
1.相互交配;孟德尔群体
2.大;小;就消失了
3.不变;仍保持不变
4.基因;基因型;隐性个体;0.8;0.2;0.64;0.04;0.32
四、名词解释
自然选择:自然界对于生物的选择作用。具有某些性状的个体对于自然界有较大的适应力从而留下较多的后代,使群体向更适应于环境的方向发展。
人工选择:人为的选择生物对人类有利的变异,并使这些变异积累和加强以形成新品种的过程。
用进废退:动物的任何器官,由于经常不断的使用,就会发达起来;相反,不经常使用,就会萎缩退化以至消失。
获得性遗传:由于环境的影响,而不是由于遗传物质的变化所形成的新的性状的遗传。
进化综合理论:是将达尔文的自然选择学说和新达尔文主义的基因论结合起来而形成的一种进化论学说。认为物种形成和生物进化的单位是种群(或称孟德尔群体),而不是单个的个体。因此,生物进化乃是群体在遗传结构上的变化。
熔解温度:使DNA/DNA或DNA/RNA双链的氢键断裂并使之解离为单链的温度。
群体遗传学:研究群体的遗传结构及其变化规律的遗传学分支学科。
孟德尔群体:指彼此间有交配关系的个体群。
基因库:一个群体中全部个体所共有的全部基因总和。
基因频率:指一个群体中不同的基因比例。
基因型频率:指一个群体中某种基因型个体数占全部个体数的比例。
哈德-魏伯格定律:在一个完全交配的群体内,如果没有其他因素(如突变、选择、迁移等)干扰时,则基因频率和基因型频率保持一定,各代不变。
突变压:由突变所造成的群体中基因频率改变的度量。
遗传漂移:由于在有限的群体内取样误差而产生的基因频率的随机波动。
地理隔离:由于地理上的障碍,而使两个或几个亲缘关系比较近的群体之间交配不易成功的隔离机制。
生态隔离:由于生活习性和栖居习性的不同,使几个亲缘关系接近的类群之间交配不易成功的隔离机制。
生殖隔离:由于生殖方面的原因使亲缘关系接近的类群之间交配不易成功的隔离机制。
物种:指形态相似,有一定分布区域,彼此可以交配,并能产生正常可育后代的一个群体。